
6. L’homéostasie et l’acclimatation
Introduction au sujet
Dans ce chapitre, nous allons faire un retour sur des notions de physiologie et voir comment on les applique à l’étude des causes et conséquences de la variation phénotypique
Objectifs spécifiques
À la fin de ce chapitre, vous serez en mesure de :
- Expliquer le concept d’homéostasie
- Décrire les composantes du centre de contrôle de l’homéostasie
- Expliquer le concept de boucle de rétrocontrôle
- Expliquer le concept d’acclimatation
- Expliquer la présence de compromis
Notions clés
1. L’homéostasie
L’environnement externe d’un organisme et son environnement interne diffèrent de plusieurs façons. Quand l’environnement externe fluctue, des processus sont enclenchés dans l’organisme pour maintenir certaines variables à l’intérieur d’une gamme de valeur. Un des concepts cruciaux de l’étude de la physiologie et du fonctionnement des systèmes physiologiques est l’homéostasie. L’homéostasie est définie comme la stabilité des systèmes physiologiques qui sont essentiels au maintien de la vie d’un organisme. Chez les animaux, on pense au pH, à la température corporelle, aux niveaux de glucose et de pression d’oxygène [1].
Lorsque la valeur d’une variable physiologique sort de son intervalle optimal, un processus s’enclenche pour la ramener vers une valeur à l’intérieur de cet intervalle. Par exemple, le niveau de glucose sanguin d’un vertébré ou la quantité de liquide extracellulaire d’une plante sont des variables physiologiques qui doivent rester dans une certaine étendue de valeur pour cette espèce (ou dans certains cas, pour cette population spécifiquement). De plus, la température corporelle d’un mammifère homéotherme est maintenue à l’intérieur d’une étendue spécifique à cette espèce, peu importe les conditions externes (figure 6.1). Dans tous ces cas, si on ne revient pas à la valeur optimale, la survie de cet organisme est carrément en péril.
Certaines espèces ont des intervalles très étroits et ne survivent que dans des conditions très spécifiques, alors que d’autres peuvent avoir une gamme étendue de valeurs face auxquelles elles survivent (par exemple, l’osmolarité de leur milieu interne), ce qui n’empêche pas qu’elles doivent maintenir l’équilibre pour d’autres variables physiologiques. Même pour un mammifère qui hiberne ou un amphibien qui estive, les conditions physiologiques durant l’hibernation ou l’estivation sont spécifiques et doivent rester à l’intérieur d’un certain intervalle. De même, pour un organisme qui migre entre des environnements très différents, comme un poisson d’eau douce qui passe une partie de son cycle vital en mer, les valeurs qui permettent de rester en vie changent avec le remodelage des systèmes physiologiques, mais la nouvelle valeur a un intervalle strict autour duquel la physiologie de l’animal doit se maintenir.
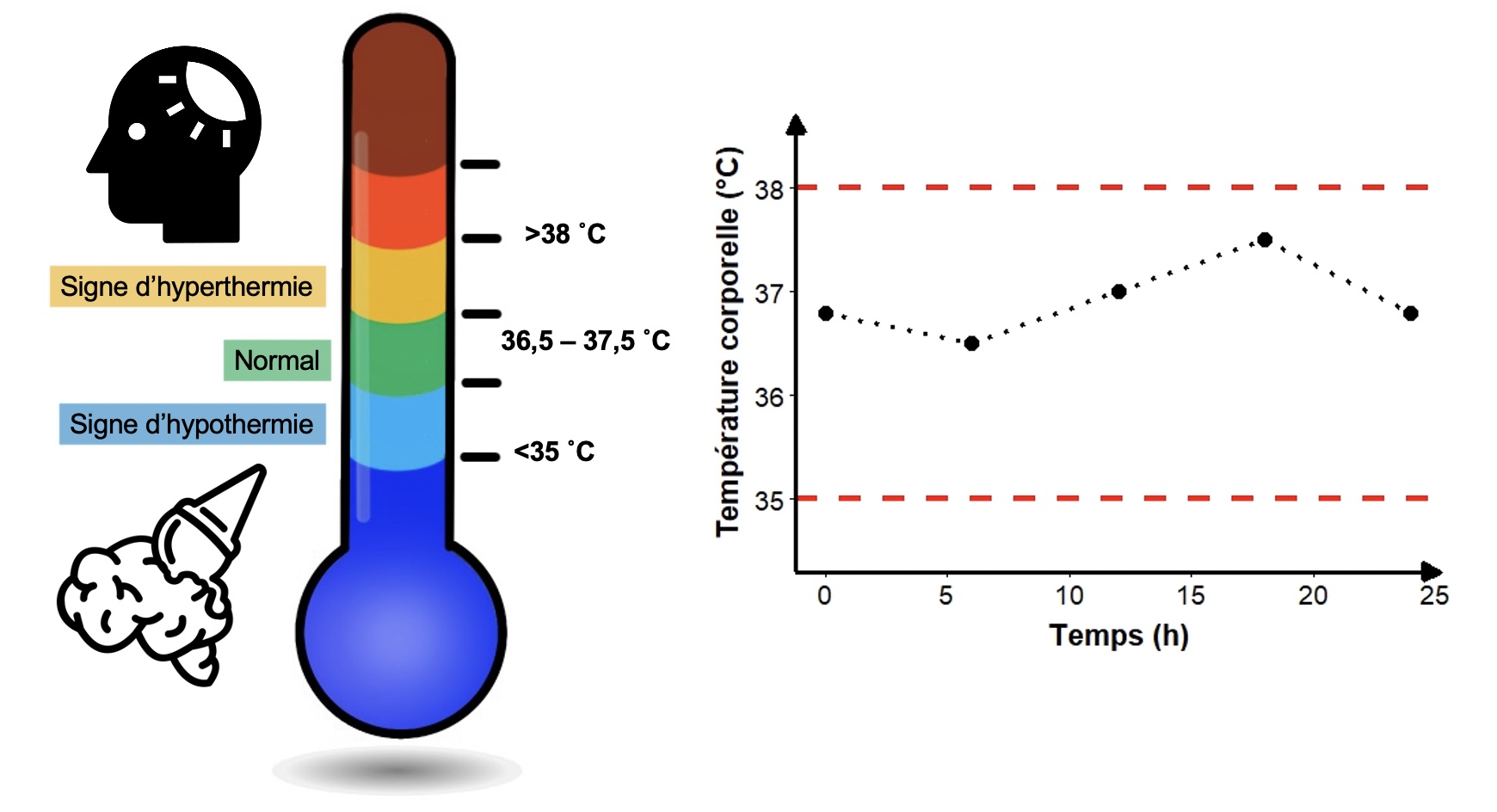
On peut donc observer en nature que, pour certains traits physiologiques, il y a une valeur moyenne dans la population et que la plupart des individus tournent autour de cette moyenne. Bien sûr, cette valeur cible peut être différente entre les espèces. On n’a qu’à penser à tous les organismes multicellulaires qui vivent en milieu marin et qui ont une gamme étendue d’osmolarité de leurs liquides intracellulaires et extracellulaires lorsqu’on compare les espèces. On trouve des espèces comme les myxines qui ont une osmolarité similaire à l’eau de mer (environ 1000 mOsm/L), et d’autres comme les poissons osseux qui ont une osmolarité autour de 250 mOsm/L. Il y a de grandes variations entre les espèces dans la valeur optimale d’osmolarité interne, mais pour une espèce donnée, il y aura très souvent une valeur optimale à maintenir pour cette variable physiologique. Bien sûr, tel que mentionné, il existe des espèces qui tolèrent de larges gammes de valeurs et d’autres des intervalles très serrés. Ce qui nous intéresse, ce sont les processus qui permettent de maintenir, et au besoin de rétablir, l’homéostasie.
2.Les composantes du centre de contrôle de l’homéostasie
Cette section est adaptée de [4].
De nombreux aspects du corps sont en constant changement : le volume et l’emplacement du flux sanguin, la vitesse à laquelle les substances sont échangées entre les cellules et l’environnement, et la vitesse à laquelle les cellules se développent et se divisent, par exemple. Ces changements contribuent en réalité à maintenir de nombreuses variables du corps à l’intérieur d’une gamme de valeurs, et donc les conditions internes globales du corps, dans des fourchettes relativement étroites. Par exemple, le flux sanguin augmentera vers un tissu lorsque ce tissu deviendra plus actif. Cela garantit que les tissus disposeront de suffisamment d’oxygène pour soutenir leur niveau de métabolisme plus élevé. Le maintien de l’homéostasie, soit la stabilité des conditions internes, qui pourrait laisser entendre qu’il ne se « passe rien », que tout est stable, repose en fait sur des processus dynamiques internes qui visent à garder la stabilité. Ces processus sont en fonction en tout temps.
Dans tous les systèmes physiologiques qui requièrent l’homéostasie, il existe donc un centre de contrôle du niveau d’une variable qui est composé d’un récepteur identifiable, d’un centre d’intégration, ainsi que d’effecteurs qui communiquent entre eux (figure 6.2). L’action de l’effecteur affecte la valeur de la variable et celle-ci retourne habituellement à l’intérieur de la gamme de valeurs attendues, ce qui est mesuré par les récepteurs et ainsi de suite. Ce centre de contrôle forme donc une boucle de rétroaction.
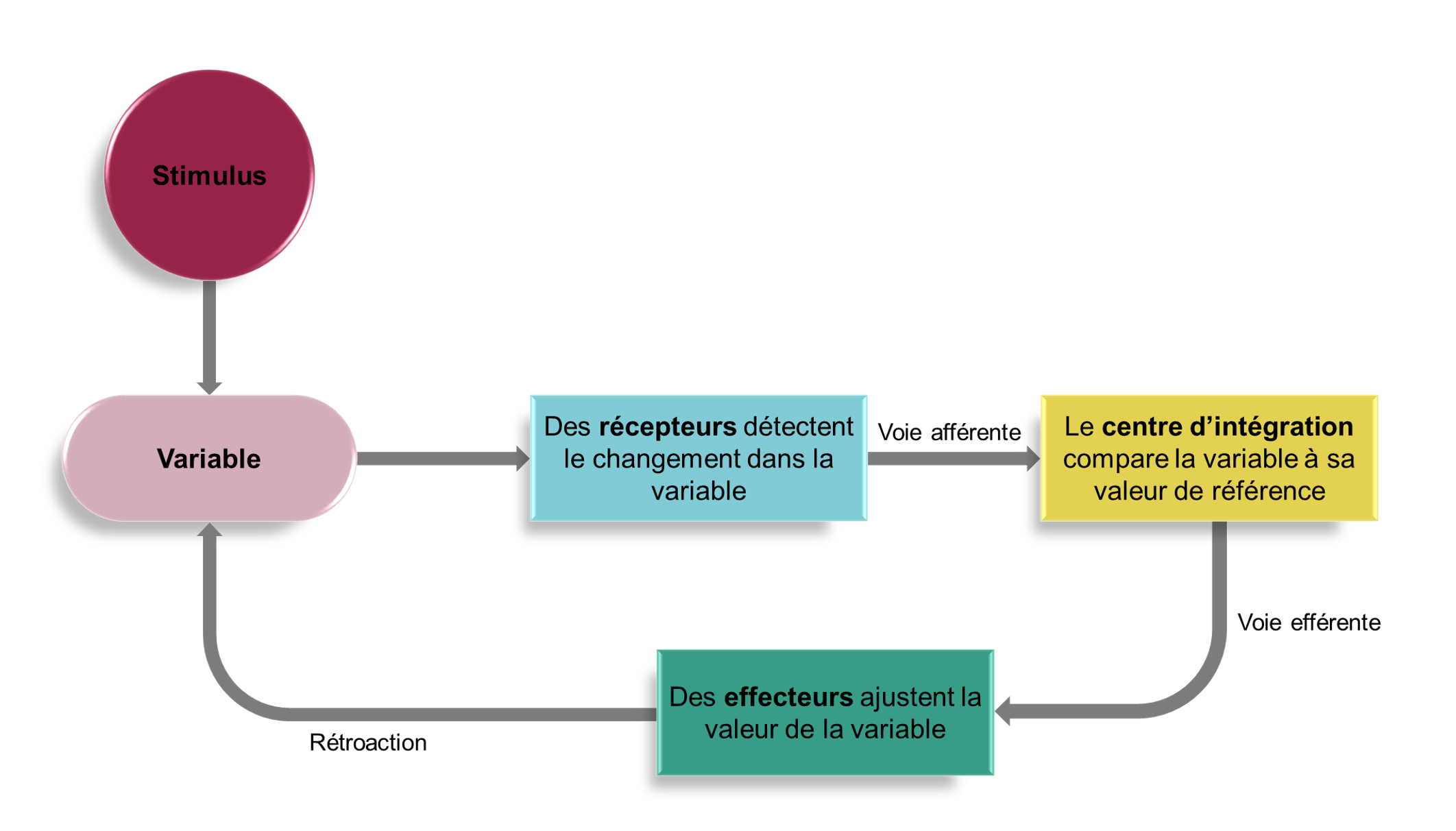
3. Les boucles de rétrocontrôle
Le maintien de l’homéostasie implique de nombreux processus régulateurs pour maintenir ou ramener à la valeur optimale une propriété de l’organisme. Ces mécanismes prennent la forme de boucles de rétrocontrôle. Les effets de ces boucles peuvent se mesurer en secondes, en minutes ou en jours.
Dans une boucle de rétrocontrôle négatif, les variables sont des paramètres surveillés et contrôlés ou affectés par le système de rétroaction. Les récepteurs mesurent la variable en tout temps et non uniquement lorsque la valeur de la variable sort de la gamme habituelle de valeurs. Ces récepteurs sont une composante essentielle car ce qui ne peut être mesuré ne peut être régulé. Chez l’humain, on peut penser aux chémorécepteurs, les barorécepteurs, les mécanorécepteurs, les thermorécepteurs, etc. Le centre de contrôle compare la variable par rapport à un point de consigne et signale aux effecteurs de générer une réponse. Les effecteurs exécutent les modifications nécessaires pour ajuster la variable. La variable change habituellement de valeur. Il faut cependant noter que l’organisme doit dépenser des ressources pour enclencher ces processus, ce qui pose une limite à la capacité de procéder à ce rétrocontrôle négatif. Des méthodes de communication entre les composantes de cette boucle de rétroaction sont nécessaires à son fonctionnement. Cela se produit souvent via les nerfs ou les hormones, mais dans certains cas, les récepteurs et les centres de contrôle sont les mêmes structures, de sorte que ces modes de signalisation ne sont pas nécessaires dans cette partie de la boucle.
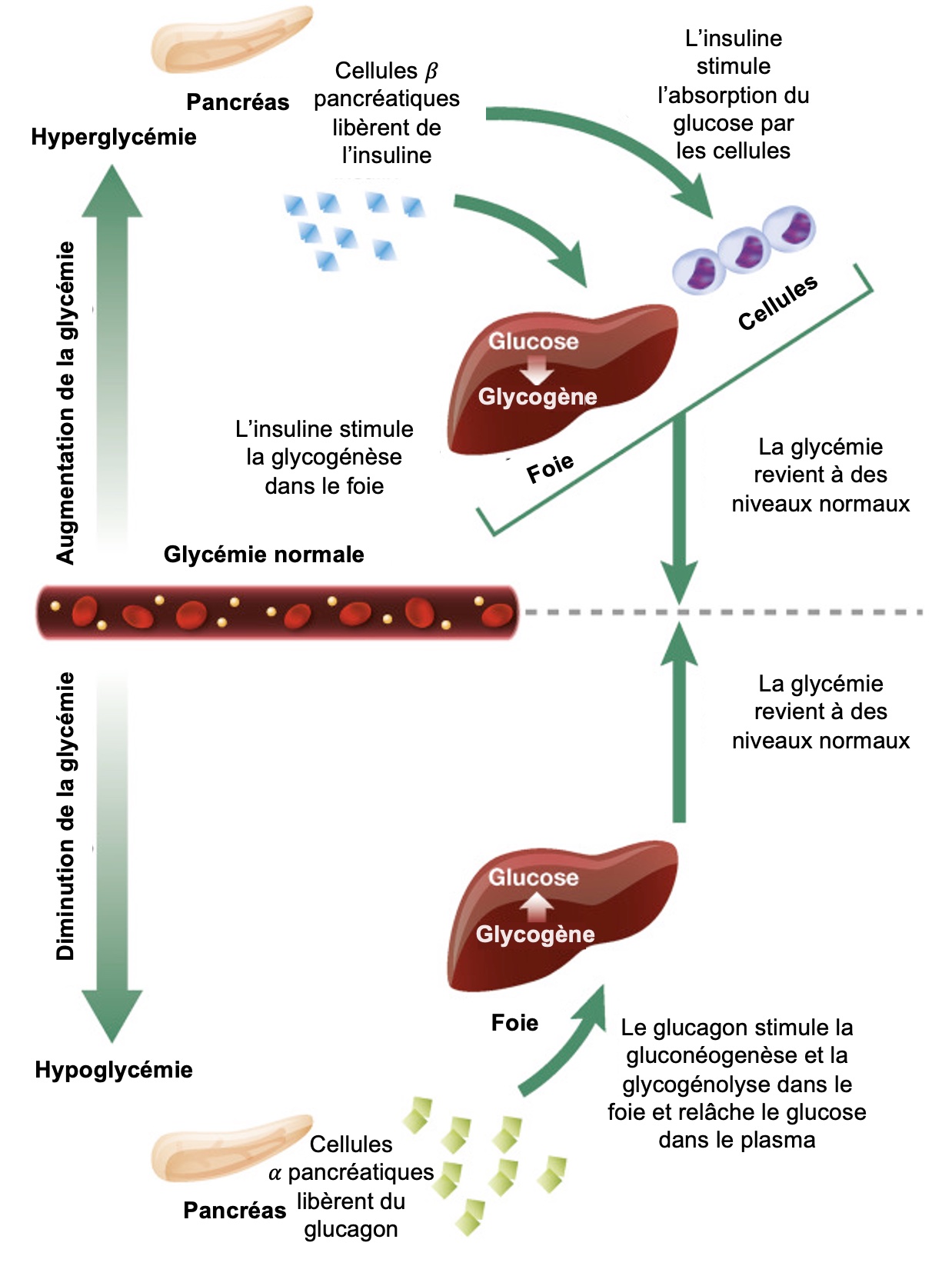
Un exemple de communication hormonale est la régulation du niveau de glucose sanguin. Si la glycémie (le taux de glucose dans le plasma sanguin) est trop basse, ceci est perçu par des récepteurs et des cellules spécialisées du pancréas sécrètent du glucagon, une hormone dont le message vise des organes qui expriment ses récepteurs, comme le foie. Le foie contient des stocks énergétiques de glycérol, qui peuvent être remis en circulation sous forme de glucose. Le glucagon a l’effet inverse de l’insuline sur les cellules (figure 6.3).
Aubin-Horth, N. (2024). Écophysiologie évolutive: les boucles de rétrocontrôle. [Vidéo]. Youtube. CC-BY. https://youtu.be/zgWL98g65uU?si=LJAylscXqi7LY5-G
Vous trouverez la transcription de l’audio de cette capsule en cliquant sur ce lien: Transcription en format word.
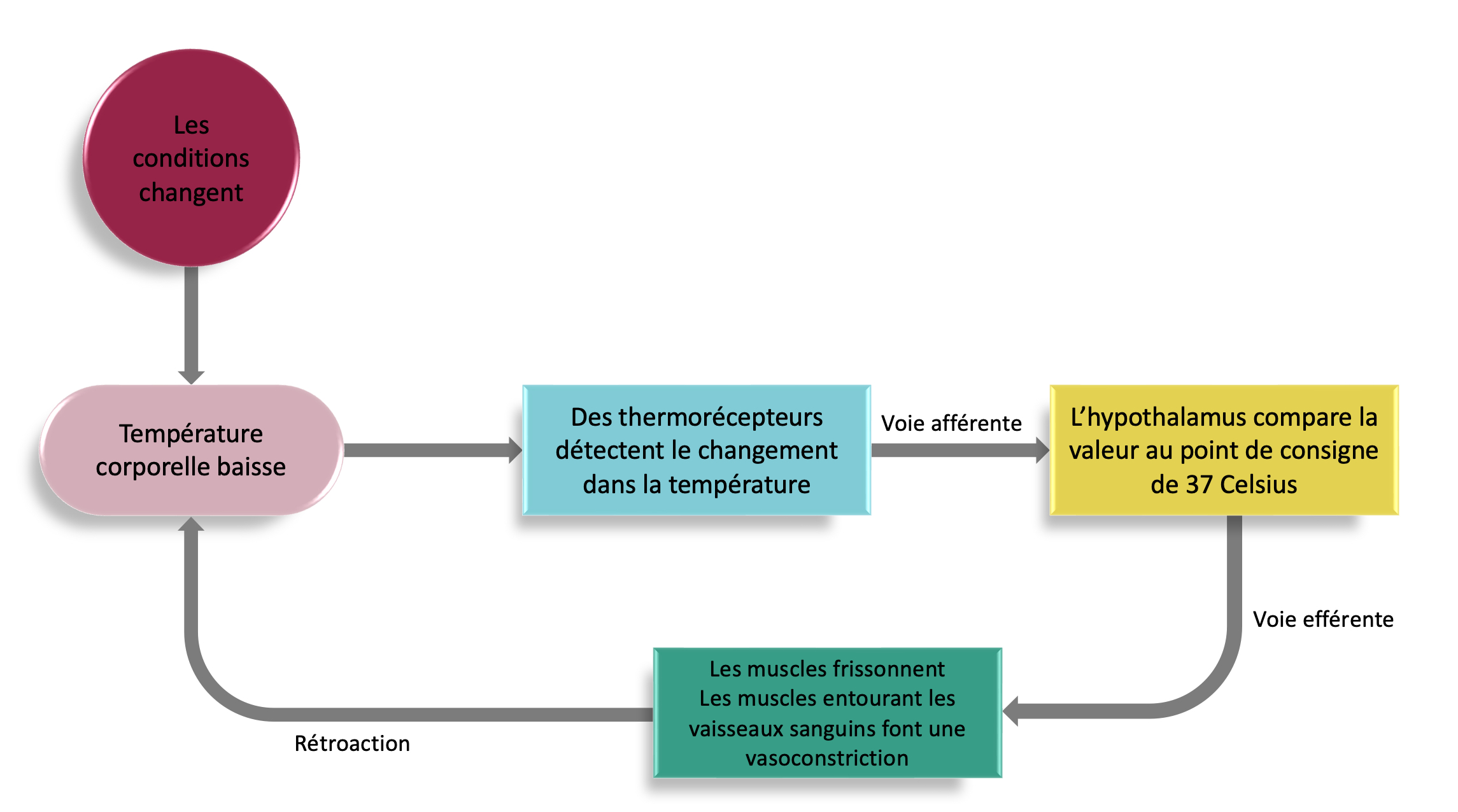
Plusieurs valeurs physiologiques sont contrôlées par les systèmes nerveux sympathique et parasympathique sans que nous ayons à réagir consciemment, telles que le maintien de la température corporelle chez l’humain (figure 6.4). Lorsque la température corporelle baisse, l’hypothalamus déclenche plusieurs réponses physiologiques pour augmenter la production de chaleur et conserver la chaleur : les frissons commencent, augmentant la production de chaleur par les muscles et les muscles entourant les vaisseaux sanguins de surface causent une vasoconstriction, ce qui diminue le flux de chaleur vers la peau. Ces effets entraînent une augmentation de la température corporelle. Lorsque la température revient à la normale, l’hypothalamus n’est plus stimulé et ces effets cessent. À l’opposé, lorsque la température corporelle augmente, l’hypothalamus déclenche plusieurs réponses physiologiques pour diminuer la production de chaleur et perdre de la chaleur : l’élargissement des vaisseaux sanguins de surface (vasodilatation) augmente le flux de chaleur vers la peau et les glandes sudoripares libèrent de l’eau (sueur) et l’évaporation refroidit la peau. Ces effets entraînent une diminution de la température corporelle. Lorsqu’il revient à la normale, l’hypothalamus n’est plus stimulé et ces effets cessent.
De nombreux mécanismes homéostatiques, comme la température et le glucose sanguin, ont des réponses différentes si la variable est supérieure ou inférieure au point de consigne. Quand la température augmente, on transpire, quand elle diminue, on frissonne. Quand le glucose sanguin augmente, on sécrète de l’insuline, lorsqu’il diminue, on sécrète du glucagon. Ces réponses utilisent différents effecteurs pour ajuster la variable. Dans d’autres cas, une boucle de rétroaction utilisera le même effecteur pour ajuster la variable vers le point de consigne, que le changement initial de la variable soit supérieur ou inférieur au point de consigne. Par exemple, le diamètre pupillaire est ajusté pour garantir qu’une quantité appropriée de lumière pénètre dans l’œil. Si la quantité de lumière est trop faible, la pupille se dilate, si elle est trop élevée, la pupille se contracte.
Les boucles de rétroaction négatives sont des systèmes intrinsèquement stables. Les boucles de rétroaction négatives, associées aux divers stimuli pouvant affecter une variable, produisent généralement une condition dans laquelle la variable oscille autour du point de consigne. On peut penser à un thermostat dans une maison qui maintient la température de la pièce autour de la température demandée sur l’afficheur en activant et éteignant un calorifère.
Ce genre de processus est couteux pour l’organisme en termes d’énergie, mais cela est contrebalancé par l’amélioration des chances de survie liée au rétablissement des systèmes physiologiques essentiels.
4. L’acclimatation
Si on mesure des valeurs physiologiques sur un individu dans différentes conditions, par exemple en comparant l’été et l’hiver, une période de sècheresse avec la saison des pluies, une photopériode longue avec une photopériode courte, ou bien un environnement en normoxie versus en hypoxie, on devrait observer des différences dans le phénotype. Ces variations dans le système physiologique d’un organisme se passent à une échelle temporelle plus longue que le maintien de l’homéostasie, mais visent aussi à maintenir et rétablir l’équilibre. C’est le processus d’acclimatation. Durant ce processus d’acclimatation, les valeurs optimales pour un système physiologique donné peuvent changer et atteindre un nouvel équilibre. Ces changements de l’environnement qui entrainent l’acclimatation peuvent être prévisibles, comme la migration, la saison de reproduction, ou un changement de conditions environnementales liées aux saisons. Ils peuvent aussi être imprévisibles, par exemple une baisse de la quantité d’oxygène en milieu aquatique.
On peut définir l’acclimatation comme un remodelage des systèmes physiologiques en réponse aux conditions de l’environnement qui vise à diminuer l’impact des nouvelles conditions de l’environnement sur l’organisme. Les changements sont réversibles et se produisent en quelques heures, jours ou semaines. L’acclimatation est un processus qui se passe face à de nombreux changements de l’environnement externe et interne, incluant la température, le pH des fluides, l’apport en eau et l’humidité ambiante, la pression partielle d’oxygène, la présence de lumière et la durée du jour, pour ne nommer que ceux-là. Les changements requis se feront dans les systèmes physiologiques, le fonctionnement cellulaire et au niveau biochimique, par exemple en modifiant la fluidité des membranes cellulaires ou en redirigeant l’énergie pour la croissance de certains organes, comme les muscles du vol.
L’acclimatation est une des causes de la variation phénotypique qu’on observe en nature. Le fait d’avoir la capacité de moduler sa physiologie face à des changements de l’environnement à moyen terme peut avoir un grand impact sur la performance et le fitness. La capacité d’acclimatation peut donc être considérée comme une adaptation. En effet, une espèce pourrait avoir la capacité de remodeler la physiologie de ses muscles entre les saisons froides et chaudes et ainsi maximiser son fitness dans chaque environnement, puis revenir à son état original au retour des conditions de départ. Au contraire, une autre espèce pourrait ne pas avoir cette capacité d’acclimatation : lorsque les conditions changent avec les saisons, les individus de cette espèce meurent.
La littérature en physiologie sur l’étude de l’acclimatation regorge d’exemples fascinants qui permettent de comprendre comment les organismes vivent dans leur habitat, mais aussi d’essayer de prédire s’ils pourront faire face aux changements rapides de l’environnement causés par les humains. Par exemple, des physiologistes peuvent mesurer la performance des individus qu’on expose à des températures de plus en plus hautes, et ce pour des groupes acclimatés à trois températures. On estime la performance d’un individu en quantifiant ses battements cardiaques. Les physiologistes quantifient l’effet de l’acclimatation préalable sur la performance en représentant graphiquement la performance à chaque température pour chaque groupe acclimaté à une température donnée.
Vérifiez votre compréhension du concept d’acclimatation avec ce quiz formatif.
Exercices
Variation génétique ou acclimatation?
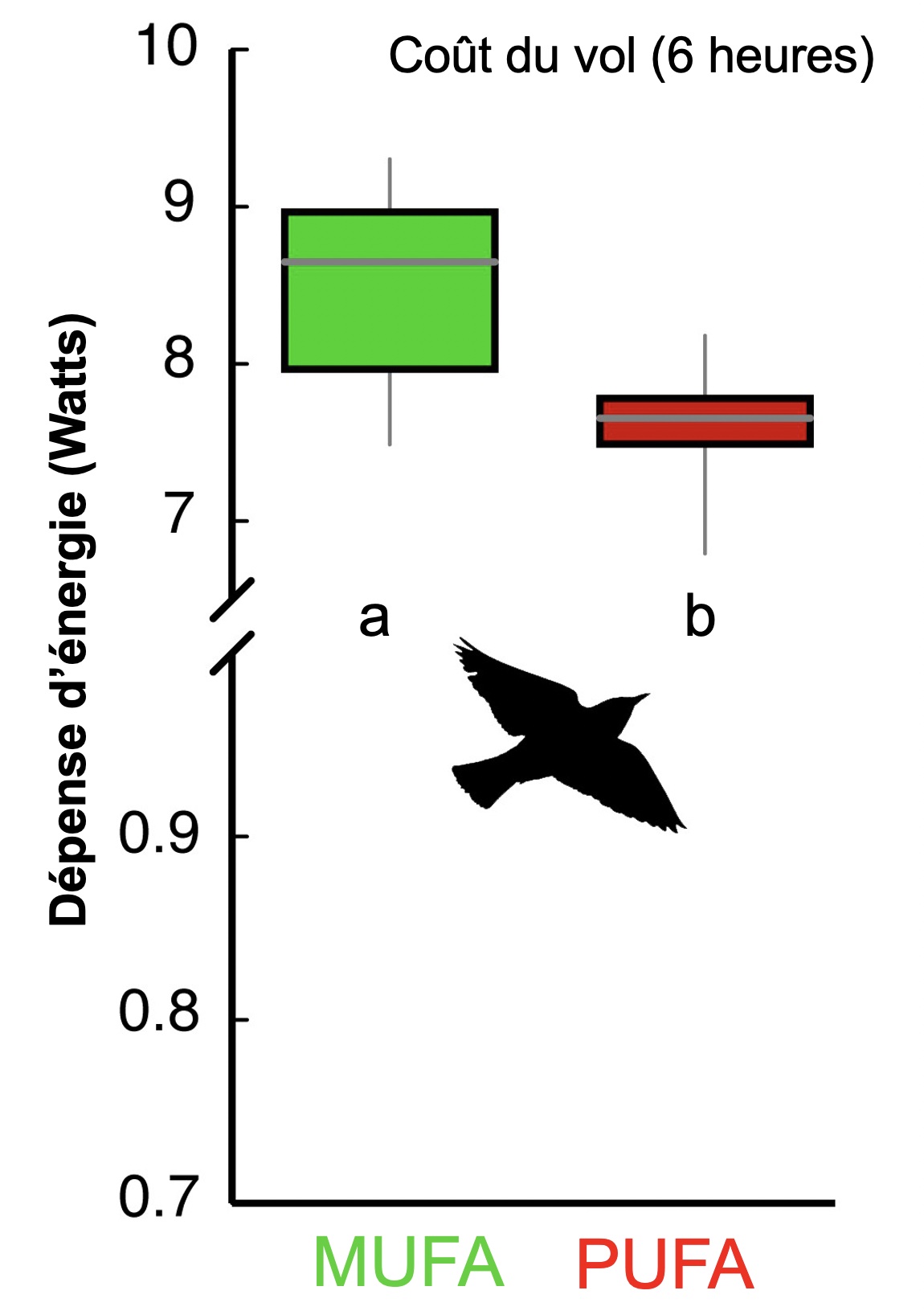
Prenons l’exemple d’étourneaux sansonnets (Sturnus vulgaris) qui se préparent à la migration. Des biologistes ont démontré que la réserve de gras qu’ils utiliseront pour le vol d’endurance a une composition en acide gras qui diffère de la composition avant la saison de migration. En effet, les individus prêts à la migration ont plus de gras polyinsaturés. L’hypothèse des biologistes était que cette acclimatation saisonnière est liée aux dépenses énergétiques du vol. Leur prédiction était que les étourneaux avec un tel profil d’acides gras polyinsaturés réduiraient leurs coûts énergétiques de vol. Afin de tester cette prédiction, les biologistes ont créé deux groupes d’oiseaux, un groupe nourri de façon à obtenir le profil d’acide gras polyinsaturé des oiseaux prêts à la migration et un autre nourri pour avoir un profil composé majoritairement de gras monoinsaturés. Lorsqu’ils ont fait voler les oiseaux des deux groupes dans un tunnel de vent pour une durée équivalente à une distance de vol de 260 km, les individus avec un profil d’acides gras polyinsaturés ont dépensé 11 % moins d’énergie que l’autre groupe (figure 6.5).
5. Les compromis
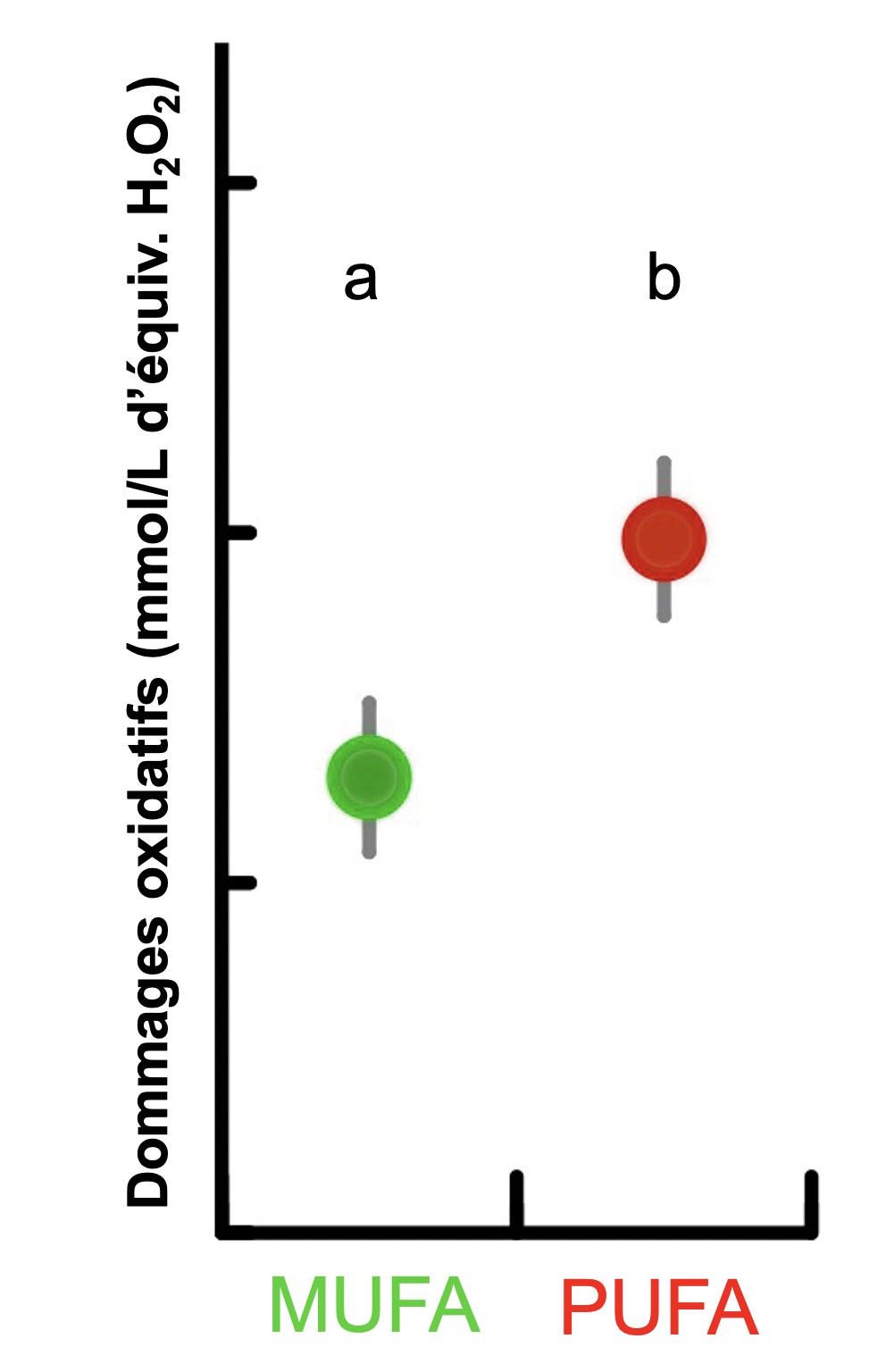
L’énergie obtenue et stockée est allouée à différents types de besoin : le maintien des fonctions quotidiennes, les changements de stade de vie comme la saison de reproduction ou la migration, ainsi que la réponse à des évènements inattendus et négatifs, comme un évènement météorologique important, la fuite face à un prédateur, ou autre. Il y a un investissement d’énergie lorsqu’un organisme met en branle des processus comme activer une boucle de rétrocontrôle pour rétablir l’homéostasie, modifier sa composition en lipides selon les saisons, ou remodeler ses cellules à la suite d’un changement de salinité. Comme la quantité d’énergie disponible pour un organisme est limitée, il y aura donc un compromis dans l’allocation d’énergie entre différents compartiments de l’organisme. Si un organisme utilise son énergie stockée pour rétablir le pH des fluides cellulaires, il ne pourra pas investir cette énergie dans la reproduction ou la croissance. De plus, certains processus sont bénéfiques à court terme mais ont des conséquences à plus long terme, tel qu’une réponse hormonale au stress ou l’utilisation de réserves de gras pour l’effort physique qui entrainera des dommages oxydatifs. Les compromis de ce type sont omniprésents en nature et sont directement reliés à l’énergie investie dans le maintien de l’homéostasie. Dans notre exemple des étourneaux sansonnets en migration présenté plus haut, les individus qui ont une composition en acide gras similaire à celle retrouvée chez les individus sauvages juste avant la migration ont une hausse significative de dommages oxydatifs (figure 6.6). Ils ont donc une meilleure efficacité de vol mais ceci est accompagné d’un coût, créant un compromis entre les deux.
Cas appliqué
L’homéostasie et les boucles de rétrocontrôle
L’homéostasie est l’état d’équilibre qui doit être maintenu pour soutenir la vie d’un organisme. Le maintien d’un niveau d’hydratation interne est essentiel au fonctionnement cellulaire et chez les organismes multicellulaires, au maintien d’un volume viable de liquides extra-cellulaires, tel que le sang. Cet équilibre est difficile à maintenir en milieu terrestre qui amène un défi important en ce qui concerne l’apport d’eau. Si le niveau d’hydratation est trop bas, un animal ressentira un signal interne que l’on nomme la soif, qui contrôle des circuits nerveux qui vont permettre de chercher une source d’hydratation (comportement), la détecter (utilisation du système sensoriel), l’utiliser (boire) et ainsi ramener l’homéostasie. La soif est donc un signal nerveux qui représente la première étape d’une boucle de rétrocontrôle qui implique plusieurs processus servant au retour à l’équilibre. Des neurobiologistes ont utilisé la drosophile (figure 6.7) pour étudier quels neurones du système nerveux central sont impliqués dans la recherche de source d’eau liée à la soif.

Les neurobiologistes ont utilisé un test de comportement où les mouches se déplacent librement et ont accès à une source d’eau (figure 6.8).

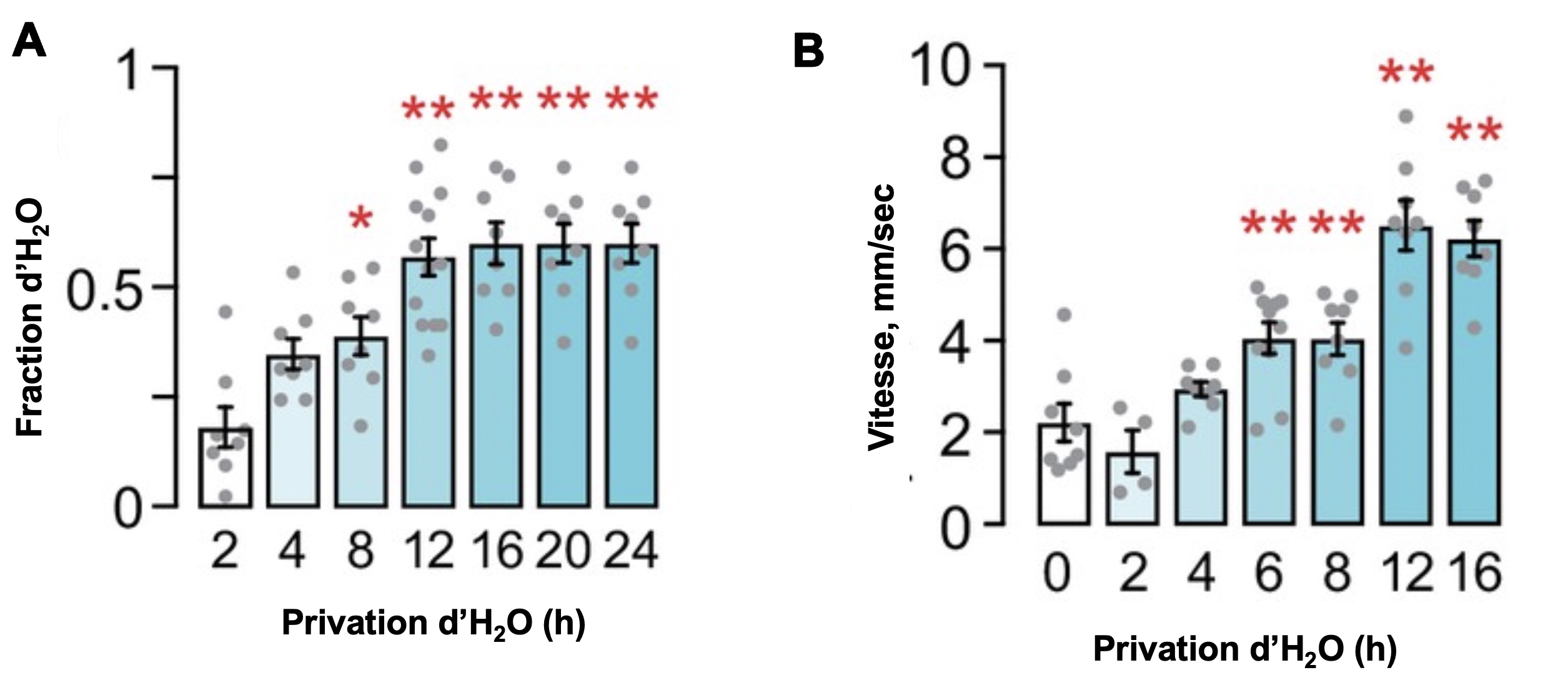
Ils ont manipulé l’homéostasie des mouches en les privant d’eau pour différentes périodes, puis ont mesuré leur comportement de recherche d’eau et la fréquence d’accès à l’eau (figure 6.9).
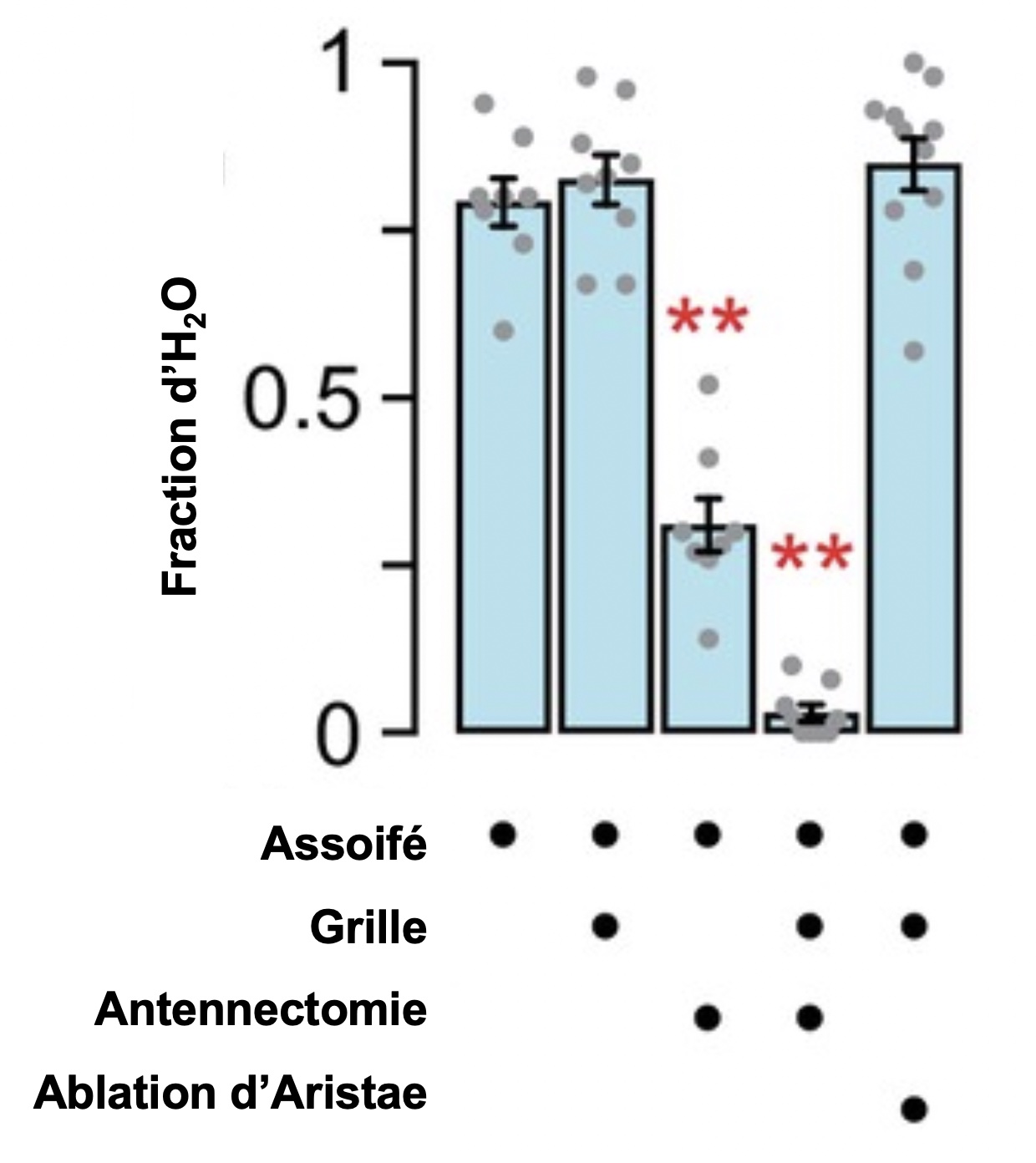
Les neurobiologistes ont testé quel segment de l’antenne est impliqué dans la détection de l’eau et son utilisation. Pour se faire, ils ont utilisé des ablations de deux parties de l’antenne. La fraction d’individus du groupe qui se retrouvent sur la source d’eau est élevée lorsque les drosophiles ont soif et lorsqu’un grillage permettant le passage de l’humidité, mais pas l’accès à la source, est installé. La fraction du groupe présente sur la source d’eau diminue fortement lorsqu’une antennectomie enlevant le troisième segment de l’antenne est réalisée. La fraction d’individus sur la source d’eau n’est pas diminuée lorsque seule une soie (appelée l’aristae) portée sur le troisième segment est enlevée (figure 6.10). Ils ont donc démontré que c’est le troisième segment de l’antenne, celui qui porte les neurones sensoriels sensibles à l’humidité, qui affecte la détection de l’eau.
Les neurobiologistes ont ensuite utilisé des lignées transgéniques de drosophile pour lesquelles des patrons neuronaux spécifiques peuvent être activés chez des mouches qui étaient en état d’équilibre hydrique, ce qui leur a permis d’isoler des neurones qui, lorsqu’activés, reproduisent la recherche d’eau, même si la mouche n’est pas assoiffée. De nombreuses expériences subséquentes avec des mutants leur ont permis de préciser exactement quels neurones enclenchent le comportement de recherche d’eau chez cette espèce.
L’acclimatation
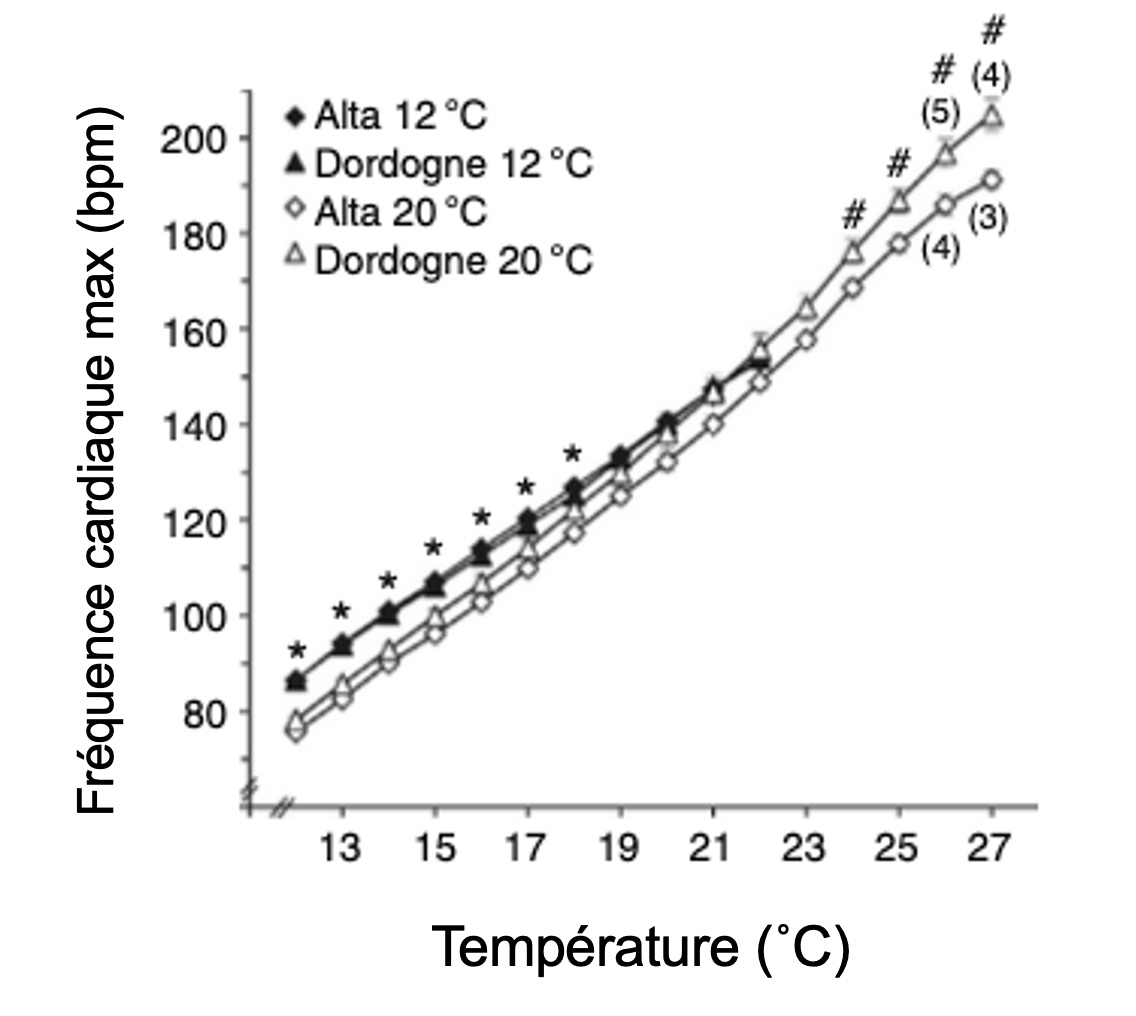
L’acclimatation est un changement réversible des systèmes physiologiques qui permet de rétablir un nouvel équilibre face à un défi de l’environnement. En physiologie, les scientifiques utilisent souvent des expériences où ils mettent des animaux face à deux ou plusieurs conditions d’acclimatation pendant plusieurs jours ou semaines pour ensuite tester leur réponse à un défi et comparer leur succès entre les traitements. Ici les physiologistes ont étudié la réponse physiologique cardiaque de saumons atlantiques juvéniles (figure 6.11) acclimatés à une température basse ou haute depuis leur éclosion, puis exposés de façon aiguë à une augmentation de la température.

Les biologistes ont élevé des juvéniles provenant de deux populations, à partir de l’éclosion jusqu’à l’âge de 3 mois dans des bassins maintenus à 12 Celsius ou à 20 Celsius. Elles ont ensuite mesuré leur rythme cardiaque en commençant à une température de 12 Celsius et en l’augmentant de 1 degré à toutes les 6 minutes. Elles ont ainsi pu quantifier le rythme cardiaque maximal, la température à laquelle les battements deviennent arythmiques et démontrer que les deux populations montrent un effet similaire de la température d’acclimatation (figure 6.12). Par exemple, la température à laquelle les saumons montrent de l’arythmie était autour de 21 à 23 C pour les individus acclimatés à 12 C, mais autour de 27,5 Celsius lorsqu’ils étaient acclimatés à 20 Celsius, montrant la capacité d’acclimatation du système cardiaque.
Résumé
L’homéostasie est un état d’équilibre. L’homéostasie est maintenue par des processus tel que des boucles de rétrocontrôle hormonales et nerveuses qui ont pour fonction de ramener l’équilibre face aux événements prévisibles et imprévisibles auxquels fait face un organisme. Ces boucles sont composées d’un récepteur, d’un centre d’intégration et d’effecteurs. L’acclimatation est un remodelage des systèmes physiologiques en réponse aux conditions de l’environnement qui vise à diminuer l’impact de ces nouvelles conditions sur l’organisme. Les changements sont réversibles et se produisent en quelques heures, jours ou semaines. Comme la quantité d’énergie disponible pour un organisme est finie, il y a un compromis dans l’investissement d’énergie entre différents compartiments de l’organisme qui fait face à des fluctuations de l’environnement.
Activité de révision
Exercices
Révision sur l’homéostasie et l’acclimatation
Bibliographie
- McEwen, B.S. et J.C. Wingfield (2003). The concept of allostasis in biology and biomedicine. Hormones and Behavior, 43(1), 2-15. https://doi.org/10.1016/s0018-506x(02)00024-7
- Demushkin, S. Sunstroke [CC-BY]. https://thenounproject.com/icon/sunstroke-412595/
- Dairy fee design. Brain freeze [CC-BY]. https://thenounproject.com/icon/brain-freeze-495204/
- Open Learning Initiative (2023) Unit 4: Homeostasis. [CC-BY-NC-SA]. https://courses.lumenlearning.com/suny-ap1/chapter/homeostasis-and-feedback-loops/.
- Hædersdal, S., A. Lund, F.K. Knop et T. Vilsbøll (2018). The role of glucagon in the pathophysiology and treatment of type 2 diabetes [CC-BY]. Mayo Clinic Proceedings, 93(2), 217-239. https://doi.org/https://doi.org/10.1016/j.mayocp.2017.12.003
- McWilliams, S., B. Pierce, A. Wittenzellner, L. Langlois, S. Engel, J.R. Speakman, O. Fatica, K. Demoranville, W. Goymann, L. Trost, A. Bryla, M. Dzialo, E. Sadowska et U. Bauchinger (2020). The energy savings-oxidative cost trade-off for migratory birds during endurance flight [CC-BY]. eLife, 9. https://doi.org/10.7554/elife.60626
- Acharya, S. (2017). A fruit fly (Drosophila melanogaster) feeding off a banana [CC-BY-SA]. https://commons.wikimedia.org/wiki/File:Drosophila_melanogaster_Proboscis.jpg
- Landayan, D., B.P. Wang, J. Zhou et F.W. Wolf (2021). Thirst interneurons that promote water seeking and limit feeding behavior in Drosophila [CC-BY]. eLife, 10. https://doi.org/10.7554/eLife.66286
- Steenstra, P. (2006). Atlantic salmon parr of Maine’s Machias River in a hatchery [domaine public]. https://commons.wikimedia.org/wiki/File:Atlantic_salmon_parr.jpg
- Anttila, K., C.S. Couturier, Ø. Øverli, A. Johnsen, G. Marthinsen, G.E. Nilsson et A.P. Farrell (2014). Atlantic salmon show capability for cardiac acclimation to warm temperatures [CC-BY]. Nature Communications, 5(1), 4252. https://doi.org/10.1038/ncomms5252

{kind=link}
{kind=link}