
5. L’étude des adaptations
Introduction au sujet
Dans ce chapitre, nous plongerons dans l’étude des adaptations, qui est bien sûr un grand thème en écophysiologie évolutive. Nous allons voir comment on définit une adaptation et ensuite nous comparerons les approches que les biologistes utilisent pour déterminer qu’un trait est effectivement une adaptation. Finalement, nous verrons comment utiliser ces différentes approches.
À la fin de ce chapitre, vous serez en mesure de :
- Définir le concept d’adaptation
- Sélectionner l’approche à utiliser afin de déterminer qu’un trait est une adaptation
- Utiliser des observations en nature pour se renseigner sur la valeur adaptative d’un trait
- Utiliser l’histoire évolutive d’un trait pour se renseigner sur la valeur adaptative d’un trait
- Utiliser la méthode expérimentale pour se renseigner sur la valeur adaptative d’un trait
- Expliquer ce qu’on peut apprendre sur les adaptations avec une expérience d’évolution expérimentale
- L’adaptation
- Comment déterminer qu’un trait est une adaptation
- Observations en nature
- Histoire évolutive d’un trait
- Évolution convergente
- Évolution divergente
- Méthodes expérimentales
- Modifier l’environnement : la transplantation
- Modifier le phénotype : l’ingénierie phénotypique
- L’évolution expérimentale
1. L’adaptation
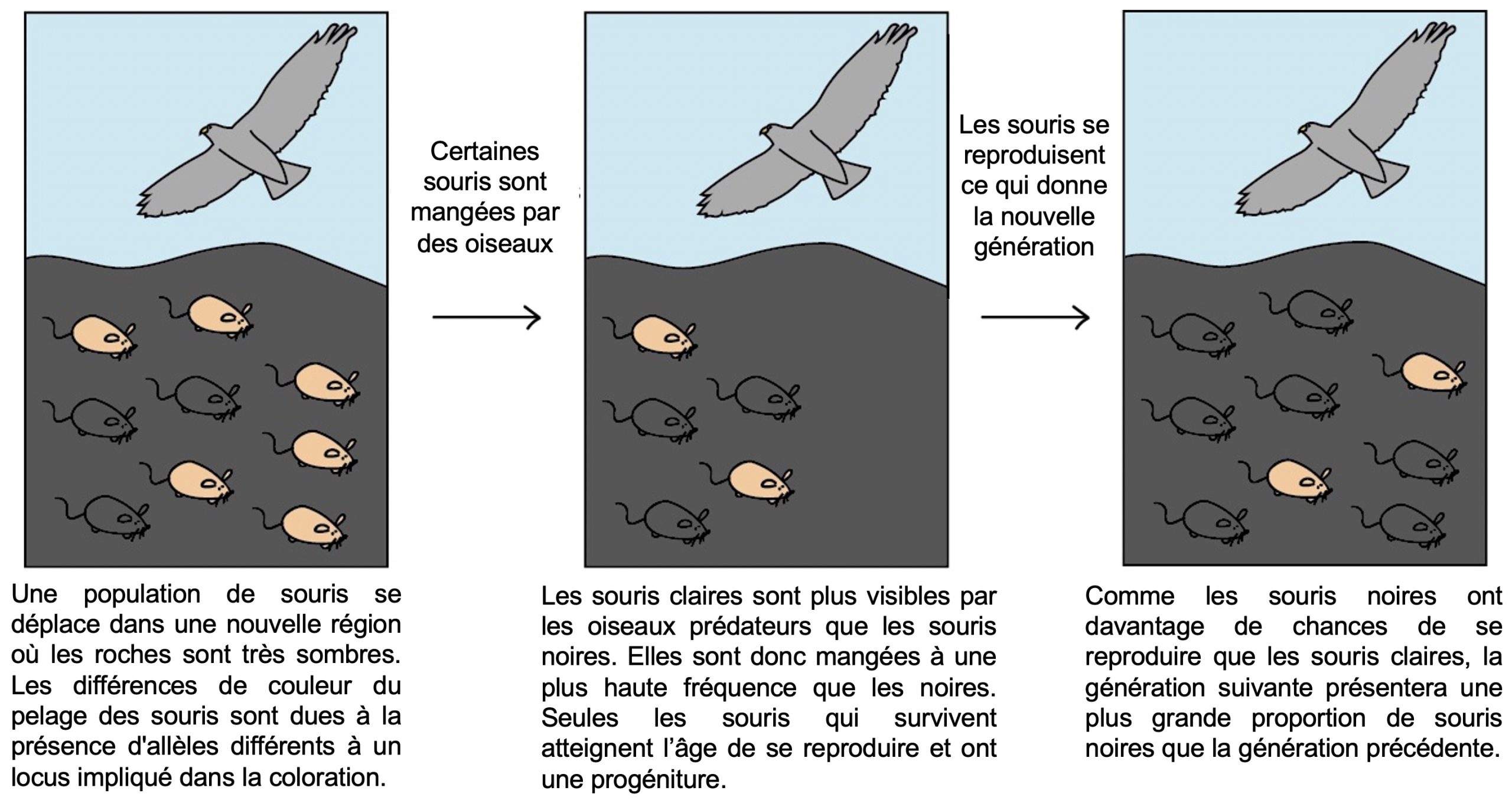
Lorsqu’on étudie un trait afin de déterminer si c’est une adaptation, la première chose à faire est de bien définir ce que l’on cherche. Une adaptation peut être définie comme un phénotype qui améliore la survie ou la reproduction de l’organisme, relativement aux valeurs alternatives de ce trait, dans un environnement donné [1]. Il est important de garder en tête qu’une adaptation est toujours évaluée « dans un environnement donné », car le même trait peut procurer un avantage pour le fitness dans un environnement, mais pas dans l’autre (figure 5.1).

2. Comment déterminer qu’un trait est une adaptation
Comme biologiste, on entend souvent parler de concept d’adaptation. Par exemple, un article scientifique peut mentionner que le trait étudié dans le projet de recherche est une adaptation. Mais comment fait-on pour savoir qu’un trait est effectivement une adaptation et que ce n’est pas seulement une déclaration basée sur une intuition? Il faut vérifier que le trait correspond bien à notre définition, c’est-à-dire que le phénotype améliore la survie ou le succès reproducteur comparé aux autres valeurs du trait dans la population, dans cet environnement. Pour y arriver, on doit combiner différents types de données.
Il y a trois approches pour déterminer qu’un trait est une adaptation. Les deux premières utilisent des observations et nous permettent d’émettre l’hypothèse que le trait est une adaptation. Ce sont les observations en nature et l’histoire évolutive d’un trait. La troisième approche nous permet de tester que le trait est une adaptation, ce sont les méthodes expérimentales.
2.1. Observations en nature
Les biologistes qui étudient un organisme peuvent faire une observation en nature et remarquer qu’un certain trait, par exemple une certaine forme de feuille, est présent chez une plante dans un habitat particulier. Cette observation en nature permet d’émettre l’hypothèse que ce trait est une adaptation. Il faudra ensuite évaluer si ce trait améliore le fitness dans cet environnement. Pour ce faire, les scientifiques devront déterminer la fonction du trait (autrement dit son effet sur la performance) et son lien avec le fitness à l’aide d’une expérience de manipulation.
Les biologistes vont aussi souvent quantifier de la variation phénotypique entre les individus. Par exemple, ils vont mesurer le phénotype de différents individus et une variable qui représente leur fitness afin de découvrir une relation entre les deux (figure 5.2). Ce type d’analyse revient à estimer la présence de sélection naturelle dans la population. Nous avons vu que pour estimer le fitness, on utilise souvent des traits qui sont directement liés à la survie et la reproduction, mais aussi parfois des traits qui « estiment » ces composantes. Par exemple, on peut mesurer le nombre d’œufs d’un poisson, la survie d’une salamandre, le nombre de fleurs d’une plante, mais aussi la vitesse de croissance d’une larve de libellule ou d’une lignée de levures, etc. Si certains individus de la population ont un meilleur fitness associé à une forme d’un trait, il sera ensuite important de déterminer quel est le lien entre cette forme du trait et le fitness. Autrement dit, on se demandera ensuite comment ce trait affecte la performance de l’organisme et bien sûr quelle est l’héritabilité de ce trait.
2.2. Histoire évolutive d’un trait
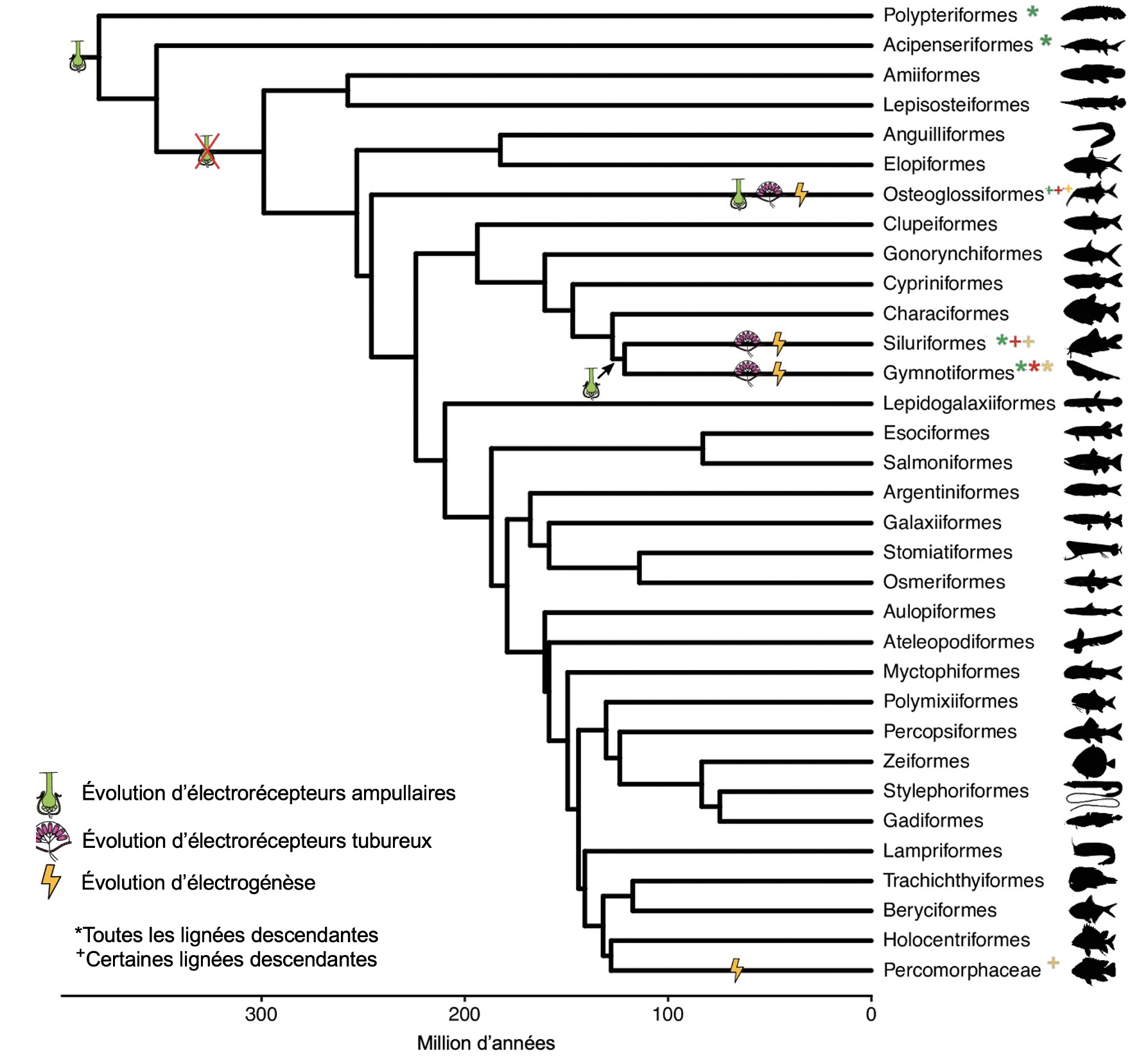
Lorsqu’on s’intéresse à l’histoire évolutive d’un trait, l’objectif est de comparer la forme ancestrale de ce trait avec la forme actuelle pour des individus qui vivent dans des habitats différents. S’il y a une association entre la forme du trait des individus et l’environnement où ils ont du succès, et que ce trait a évolué dans cet environnement, cela nous permet d’émettre l’hypothèse que c’est une adaptation. C’est une autre forme d’observation. Pour esquisser l’histoire évolutive du trait, il faut déterminer la forme du trait chez l’ancêtre de l’organisme qui nous intéresse en utilisant une analyse phylogénétique (figure 5.3). On peut estimer quand le trait est apparu durant l’évolution et déterminer la probabilité qu’il soit présent sous cette forme dans un ancêtre commun. On peut aussi déterminer quand un trait est disparu durant l’évolution de ce groupe. Ce genre d’information phylogénétique nous permettra de déterminer s’il y a de l’évolution convergente ou de l’évolution divergente.
2.2.1. Évolution convergente
L’évolution convergente est définie comme l’évolution indépendante du trait dans deux taxons ou plus. Si un trait évolue plus d’une fois de façon indépendante quand les populations font face aux mêmes défis environnementaux, cela suggère que le trait est une adaptation à ce défi de l’environnement. On observe chez différents taxons non directement apparentés des « solutions » qui se ressemblent face aux mêmes défis de l’environnement.
On peut confirmer l’évolution convergente si l’ancêtre commun de ces taxons (plus ou moins éloignés évolutivement) n’avait pas cette forme du trait. Autrement dit, l’origine du trait n’est pas unique. Nous pourrions par exemple étudier les différentes espèces d’animaux qui vivent en altitude et découvrir qu’ils ont des caractéristiques physiologiques qu’on ne retrouve pas dans les populations qui ne font jamais face à l’hypoxie, qui est un défi quotidien en altitude. Ici on verrait l’évolution répétée de traits similaires face au défi de l’environnement en altitude. Ce serait de l’évolution convergente.
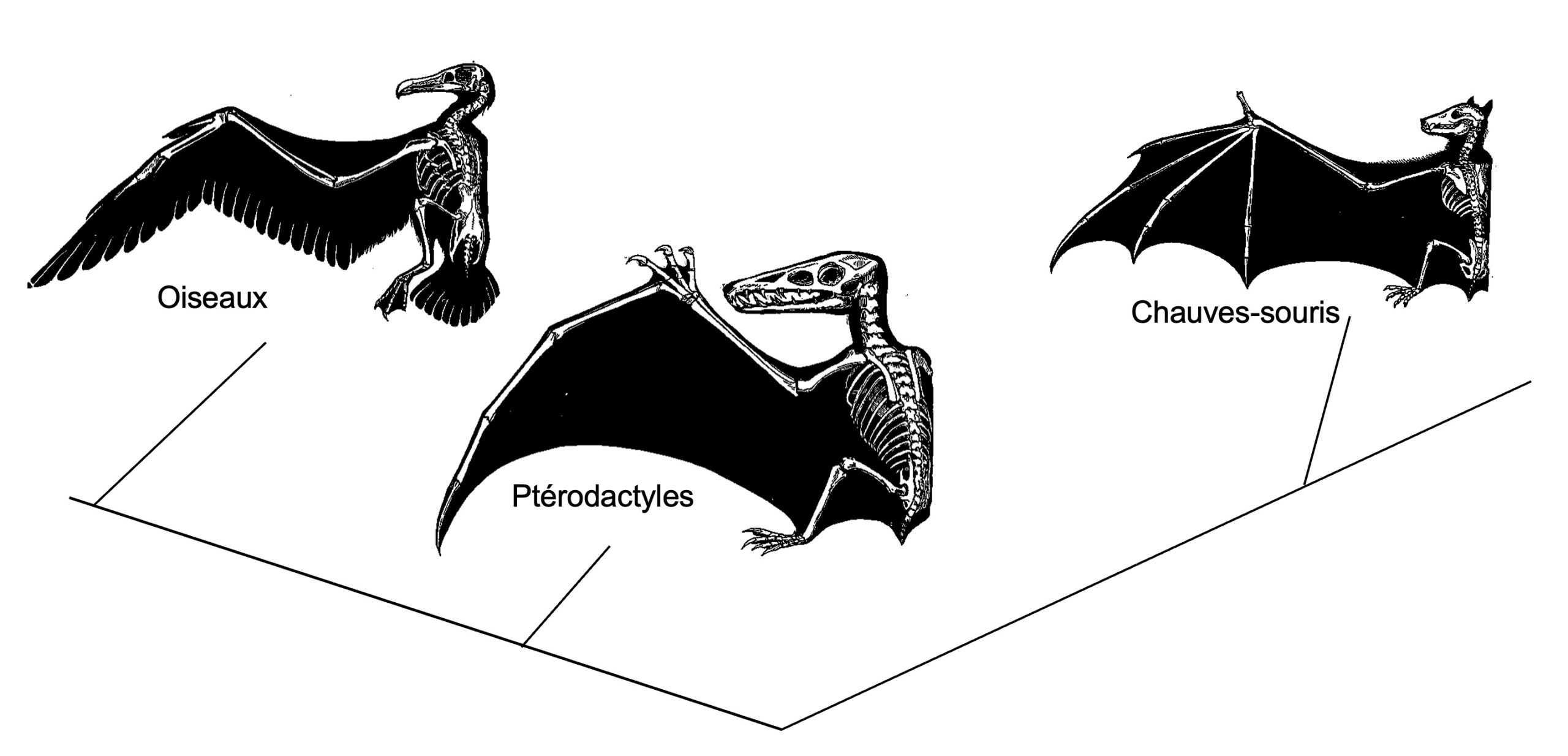
Un exemple classique d’évolution convergente sur une échelle évolutive très grande est l’apparition d’organes respiratoires internes dans les groupes d’animaux qui ont envahi le milieu terrestre comme les escargots chez les mollusques, les insectes chez les arthropodes et les humains chez les vertébrés. On peut aussi penser à l’évolution de membres en forme d’ailes utilisés pour le vol chez différents groupes non apparentés (figure 5.4). L’évolution répétée de l’électrodétection et de l’électrogénèse chez les vertébrés en est aussi un exemple (figure 5.3 plus haut), ainsi que la résistance de certains végétaux à la présence de grande quantité de fer dans le sol, ou le style de vie fouisseur accompagné de changements dans la myoglobine des muscles chez l’ordre des Eulipotyphles qui comprend des espèces de taupes, de musaraignes et de hérissons.
2.2.2. Évolution divergente
Lorsque nous analysons l’histoire évolutive d’un trait, nous pourrions au contraire détecter que des espèces très proches évolutivement ont divergé dans un trait particulier alors qu’elles utilisent des environnements différents (figure 5.5). Pour un trait physiologique, on pourrait penser à diverses capacités de digestion pour des populations de la même espèce qui n’ont pas accès aux mêmes sources de nourriture. Ici on verrait de l’évolution divergente, car ces espèces ou populations proches évolutivement ont des traits différents quand elles font face à un environnement différent.

On peut penser à un cas où ce sont les humains qui ont changé l’environnement. Les moustiques sont des vecteurs de nombreuses maladies et on dit même que c’est l’animal qui fait le plus de victimes humaines sur terre. Les humains utilisent différents produits chimiques pour tenter d’éliminer les populations de moustiques. Dans les zones où on utilise un insecticide (le DDT), les moustiques ont évolué une résistance à ce produit. Les populations de moustiques à proximité, mais qui ne sont pas en contact avec le DDT, n’ont pas divergé évolutivement. Les moustiques résistants à l’insecticide ont une forme plus performante de l’enzyme qui dégrade le DDT en substance non-toxique et cette meilleure performance augmente leur survie. Les différentes populations ont un ancêtre commun proche, mais elles sont divergentes pour un trait, ce qui permet d’émettre l’hypothèse que ce trait est une adaptation.
Aubin-Horth, N. (2024). Écophysiologie évolutive: Comment déterminer qu’un trait est une adaptation – partie 1. [Vidéo]. Youtube. CC-BY. https://youtu.be/0oqjxGvrIew?si=RM1xlyjvm_uwwm7L
Vous trouverez la transcription de l’audio de cette capsule en cliquant sur ce lien: Transcription en format word
2.3. Méthodes expérimentales
On vient de voir qu’on peut faire des observations en nature ou utiliser l’histoire évolutive d’un trait pour émettre l’hypothèse que ce trait là est une adaptation. Pour pouvoir tester les prédictions qui découlent de cette hypothèse, on doit utiliser des manipulations expérimentales. On peut utiliser la définition d’une adaptation pour se donner une idée de ce que nous devrons manipuler!
Basé sur notre définition de l’adaptation, notre prédiction pourrait être que si on change l’environnement tout en conservant la valeur du trait, le trait sera associé à un fitness moins élevé. Pour tester cette prédiction, nous pouvons faire une manipulation expérimentale dans laquelle on change l’environnement et on mesure le fitness du même trait dans de nouvelles conditions environnementales. On parle alors d’une expérience de transplantation.
Nous pourrions aussi émettre la prédiction qu’une autre valeur de ce trait serait associée à un fitness plus faible si on conserve l’individu dans cet environnement. Notre approche dans ce cas-ci est de changer la valeur du trait et de mesurer le fitness dans le même environnement. On parlera alors d’ingénierie phénotypique.
Finalement, on peut manipuler l’évolution elle-même et quantifier ce qui change dans les traits de l’organisme qui lui permettent de faire face au défi de l’environnement qu’on a créé. C’est l’évolution expérimentale.
2.3.1. Modifier l’environnement : la transplantation
Dans une expérience de transplantation, nous gardons la valeur du trait constante et nous manipulons l’environnement. Le fitness des individus est mesuré dans chaque environnement : original et environnement manipulé. C’est pour ça qu’on parle de transplantation; c’est comme si on transplantait l’individu dans un autre environnement. Étant donné notre hypothèse que ce trait est une adaptation, notre prédiction est que le fitness sera moins élevé dans le nouvel environnement. Si on compare deux populations qui ont des traits différents, on peut chacune les élever dans leur environnement d’origine et dans l’autre environnement. On parlera alors dans la littérature de transplantation réciproque.
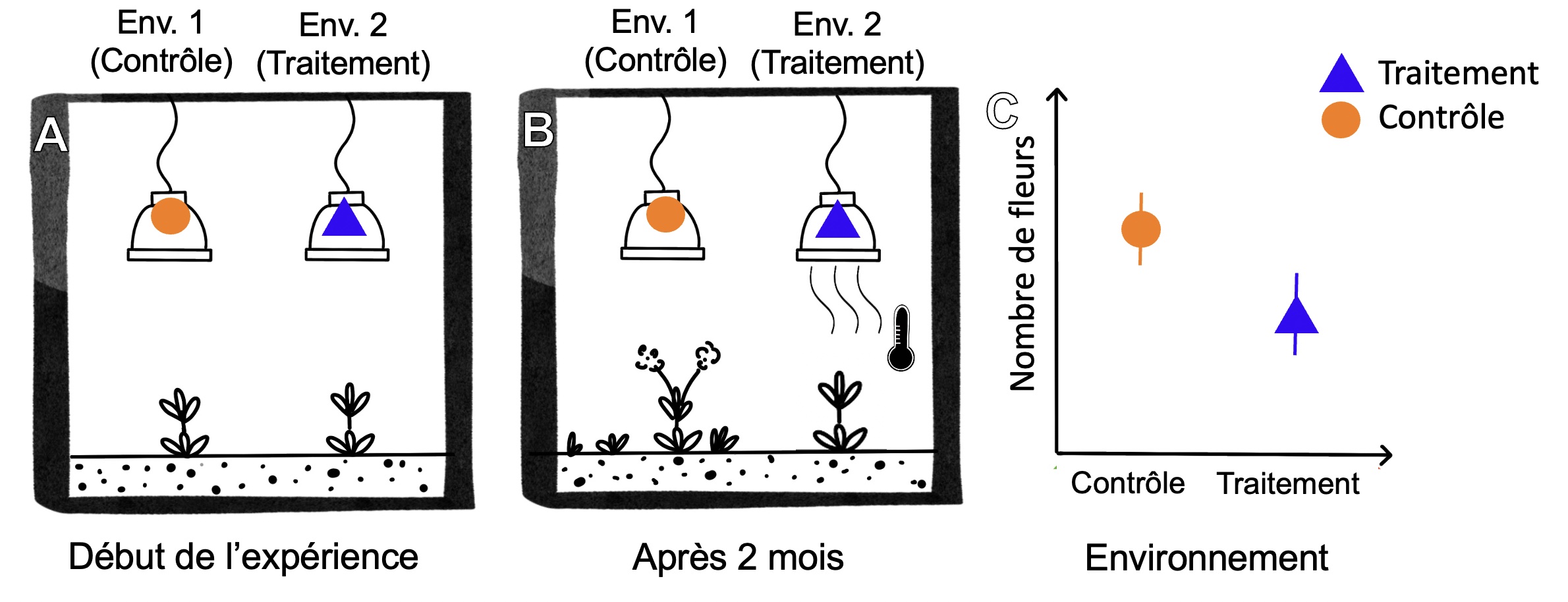
Pour manipuler l’environnement, il faut avoir une idée de l’agent de sélection. Par exemple, doit-on manipuler la présence de prédateurs, la température de l’eau, la quantité d’oxygène? Si on prend l’exemple fictif d’une plante qui vit en milieu tempéré et qui semble adaptée à son environnement. Si on créé un traitement où la température est élevée, on peut mesurer les effets de transplanter cette plante sur le fitness et tester notre prédiction (figure 5.6).
2.3.2. Modifier le phénotype : l’ingénierie phénotypique
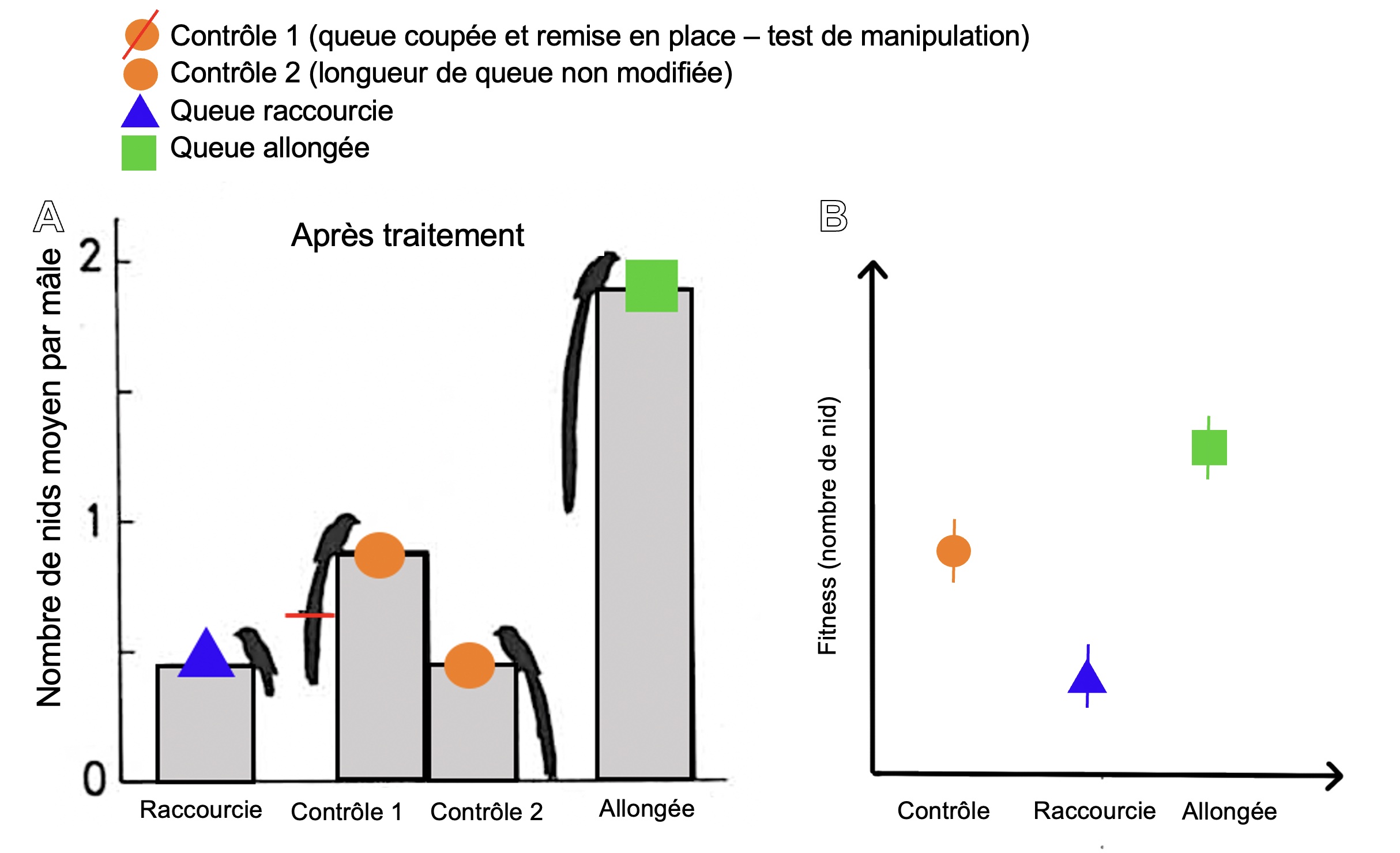
Dans une expérience d’ingénierie phénotypique, nous gardons l’individu dans son environnement, mais nous manipulons la valeur du trait qui nous intéresse. On mesure ensuite le fitness du trait avec sa valeur originale et lorsqu’elle est manipulée, dans le même environnement. Comme notre hypothèse est que le trait original « améliore la survie ou la reproduction de l’organisme, relativement aux valeurs alternatives de ce trait, dans un environnement donné », soit la définition d’une adaptation, notre prédiction est que le fitness sera plus bas pour le trait modifié dans cet environnement.
Pour modifier la valeur du trait, nous pouvons faire une manipulation physique qui affecte l’apparence de l’organisme (figure 5.7). Nous pourrions aussi faire une manipulation pharmacologique avec une drogue qui bloque l’effet d’une hormone ou qui augmente son niveau en circulation, par exemple. Finalement, nous pourrions faire une manipulation génétique : utilisation d’ARN interférent (« RNAi) pour rendre un gène silencieux, édition de gène avec des approches comme CRISPR-Cas9, etc.
Dans tous les cas que nous avons vus jusqu’à présent, nous avons identifié les variations dans un trait et le lien avec l’environnement ou le fitness, entre des espèces, entre des populations, ou à l’intérieur d’une population. Cependant, comment peut-on quantifier l’effet de la valeur d’un trait sur la performance et le fitness si tous les individus ont la même valeur de trait dans cet environnement? Par exemple, pensez à un trait physiologique qui aurait une valeur très similaire entre les individus d’une population, comme la capacité chez les poissons qui vivent en eau salée de transporter des ions par les cellules de leurs branchies, ce qui leur permet de contrôler l’osmolarité interne qui est différente de celle du milieu ambiant. Comment faire pour savoir si ce trait est une adaptation si tous les individus ont la même capacité? Dans un cas comme celui-ci, on peut avoir recours à l’ingénierie phénotypique pour « créer » de la variation phénotypique, ce qui nous permettra de tester des prédictions sur la valeur adaptative du phénotype!
Exercices
Vérifiez votre compréhension des différentes manières de déterminer si un trait est une adaptation avec ce quiz formatif.
Associer les différentes manières de déterminer si un trait est une adaptation en utilisant l’exemple des pinsons de Darwin.
2.4. L’évolution expérimentale
Une approche complètement différente pour étudier la cible de la sélection et les bases génétiques d’une adaptation est de créer expérimentalement l’agent de sélection et de mesurer quels traits changent (ou non) face à cette pression de sélection (figure 5.8). Cela nous indique quelle est la cible de la sélection et quels sont les changements génétiques et moléculaires associés.
Par exemple, on peut étudier des drosophiles qu’on expose à la sècheresse sur plusieurs générations en laboratoire, en maintenant une lignée témoin pour la même durée. On étudie quels systèmes physiologiques sont altérés chez les lignées qui font face à la sècheresse et on mesure le fitness dans l’environnement original et dans l’environnement expérimental pour les deux lignées. On peut ainsi voir si la modification de certains traits au cours de l’évolution expérimentale est associée à une amélioration du fitness comparé au trait original et déterminer quels sont les changements génétiques qui sont associés. Est-ce qu’il y a une mutation dans la séquence codante d’un gène qui modifie une enzyme? Est-ce que c’est une mutation dans la séquence régulatrice d’un gène qui code pour un canal ionique qui change la quantité de ce canal dans la membrane? Etc.
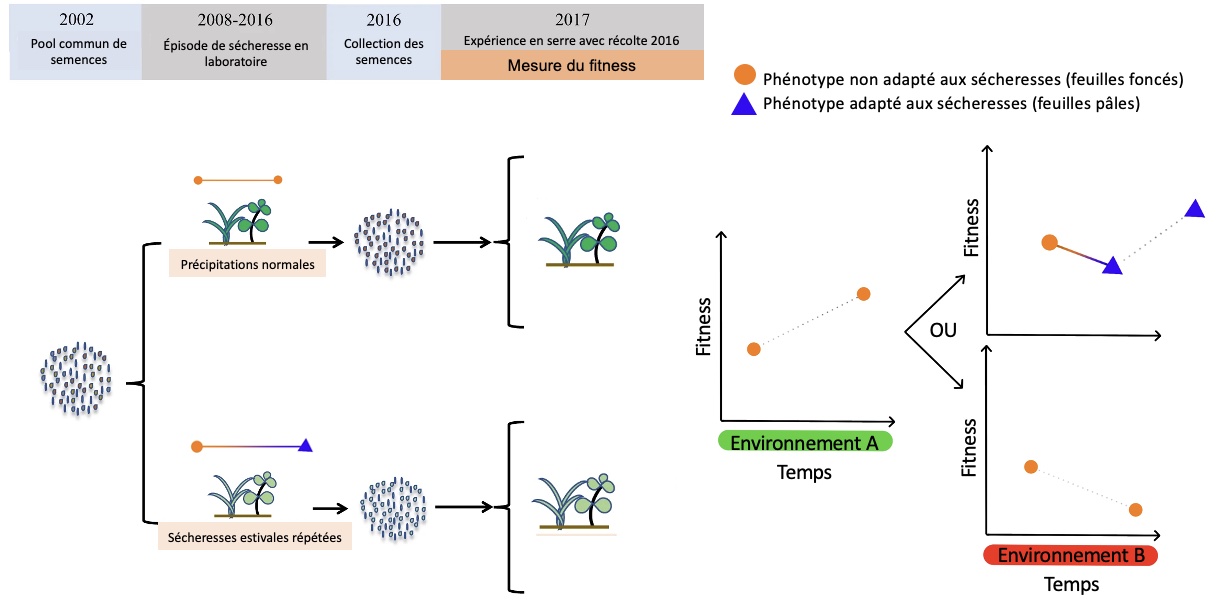
Si on répète l’expérience d’évolution expérimentale plusieurs fois en utilisant des génotypes différents comme populations de départ (drosophiles d’une population au nord, ou vivant en altitude, ou en milieu humide, etc.), on peut tester si tous ces génotypes auront la même trajectoire évolutive et si les mêmes systèmes physiologiques seront la cible de la sélection. L’évolution expérimentale est un outil puissant en biologie évolutive.
Aubin-Horth, N. (2024). Écophysiologie évolutive: Comment déterminer qu’un trait est une adaptation – partie 2. [Vidéo]. Youtube. CC-BY. https://youtu.be/zQnvPhyTunA?si=9mbLmSh9LNF29ka9
Vous trouverez la transcription de l’audio de cette capsule en cliquant sur ce lien: Transcription en format word
Cas appliqué
Entrevue avec Markus Thormeyer
Cette capsule vidéo présente une entrevue avec Markus Thormeyer, étudiant au doctorat en zoologie dans le laboratoire de Michelle Tseng, à UBC. Ses travaux portent sur l’évolution des moustiques en milieu urbain, particulièrement face aux ilots de chaleur, comparativement aux milieux ruraux. Vous pouvez activer les sous-titres en français ou en anglais.
Aubin-Horth, N. (2023). Écophysiologie évolutive: Markus Thormeyer. [Vidéo]. Youtube. CC-BY. https://youtu.be/T7aaul-i7oQ?si=kUbvy5sNQ2L8Fu7Z
Vous trouverez la transcription de l’audio de cette capsule en cliquant sur ce lien. Transcription Thormeyer en format word
Histoire évolutive d’un trait : l’évolution convergente
Des biologistes ont étudié la viviparité chez des lézards de la famille des Phrynosomatidés (figure 5.9). Il se sont demandé si l’évolution répétée de la viviparité est associée à des changements aussi répétés dans la physiologie, comme la température critique minimale et maximale (CT min et CT max).

Les physiologistes ont étudié une famille de lézards qui sont largement vivipares et chez qui il y a eu une transition vers la viviparité à cinq reprises durant l’évolution (figure 5.10). Les ancêtres représentés en rouge dans la figure n’avaient pas ce trait alors que certains descendants sont vivipares (en bleu). Cette transition à cinq reprises vers la viviparité de façon indépendante représente un cas d’évolution convergente.
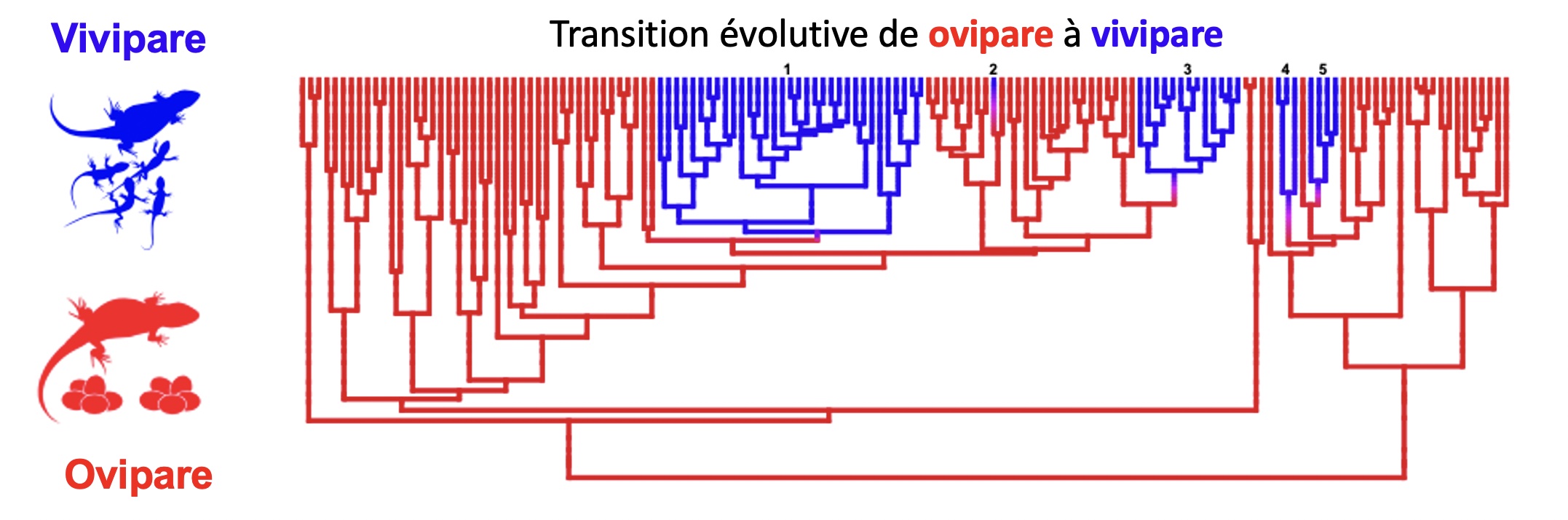
Les physiologistes ont émis l’hypothèse que ce changement d’histoire de vie est associé à l’utilisation d’habitats plus froids. Ils ont prédit que si c’est le cas, des traits physiologiques associés à la vie dans un habitat plus froid sont aussi apparus de façon répétée. Ils ont trouvé que les lézards vivipares ont en effet une température critique minimale et maximale plus basse et que la température de leur corps est plus basse de 4 degrés Celsius en moyenne, suggérant une évolution convergente de la physiologie thermique chez cette famille de reptiles.
Méthodes expérimentales : Modifier l’environnement avec la transplantation
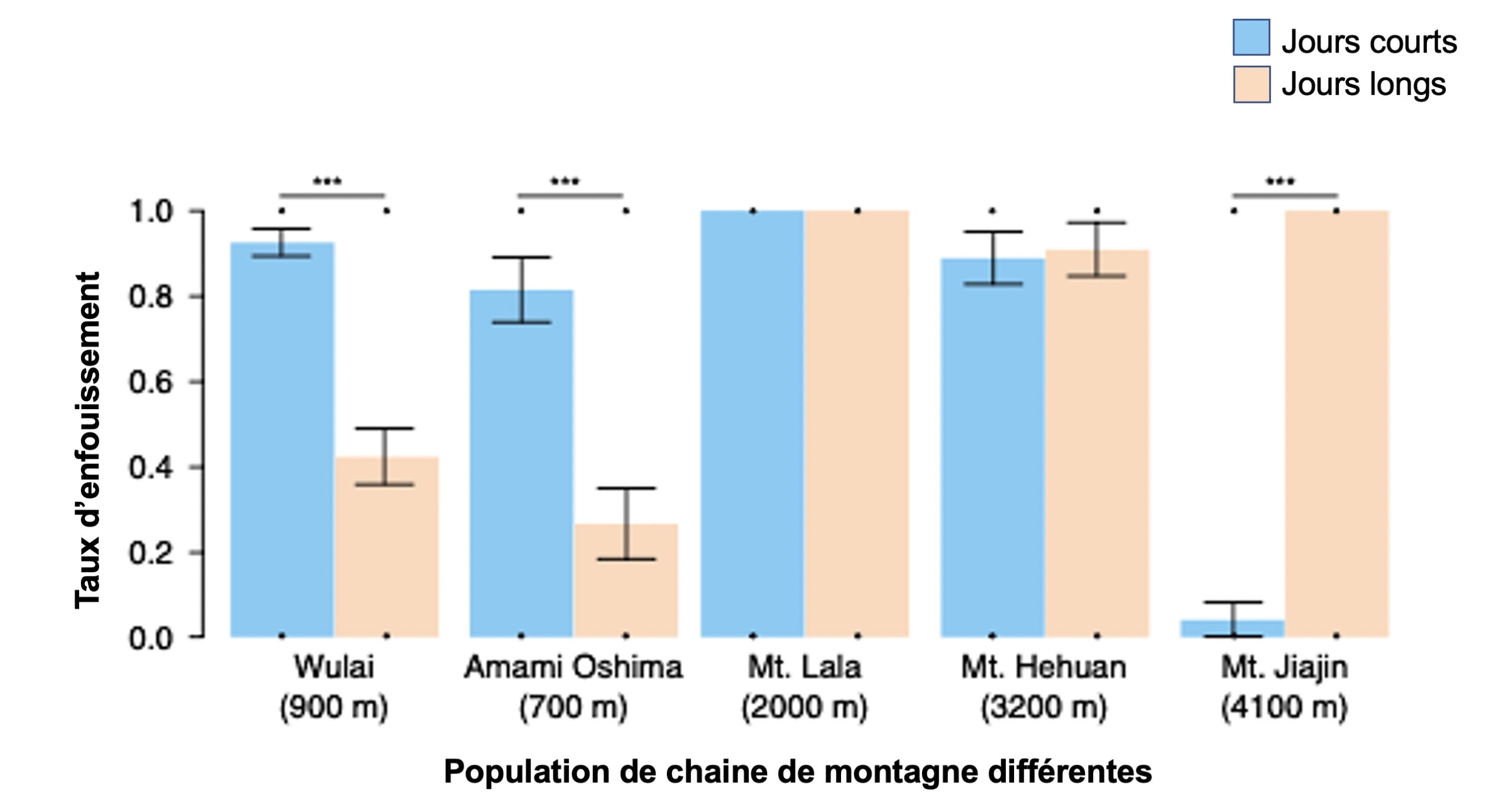
Des biologistes s’intéressent à la reproduction chez les insectes. Ils ont étudié différentes populations d’un coléoptère nécrophore, Nicrophorus nepalensis, qui enterre des carcasses d’animaux pour nourrir ses rejetons (figure 5.11).

Les scientifiques veulent savoir si ces populations qui vivent à différentes altitudes dans les montagnes en Asie ont une reproduction contrôlée par la photopériode et si oui, si cela est une adaptation locale aux conditions très différentes entre les habitats. Elles ont utilisé une expérience de jardin commun pour tester l’effet de la photopériode sur le comportement de reproduction dans 5 populations de cet insecte. Elles ont placé des individus sous une photopériode à jours courts (ce qui correspond à l’hiver à nos latitudes) et à jour long (l’été) et ont mesuré le taux d’enfouissement de cadavres, une mesure de l’activité de reproduction. Elles ont observé des patrons très variés (figure 5.12) : deux populations se reproduisaient significativement plus dans le traitement à jour court, suggérant qu’elles se reproduisent l’hiver (ex : Wulai qui se situe à 900 mètres d’altitude), deux autres avaient une reproduction élevée sous toutes les photopériodes, suggérant qu’elles se reproduisent toute l’année et que la photopériode ne les affecte pas (ex : Mt. Hehuan à 3200 m) et la dernière, retrouvée au Japon à 4100 m, se reproduisait seulement sous la photopériode de jours longs, donc l’été.
Les scientifiques ont ensuite utilisé une expérience de transplantation réciproque pour mesurer l’effet sur le fitness de ces différences de réponse à la photopériode. Elles ont mesuré le succès de reproduction d’individus venant des populations de Wulai (reproduction hivernale) et du Mont Hehuan (reproduction tout l’année indépendante de la photopériode) lorsque transplantées à Wulai ou au Mont Hehuan en été ou en hiver (figure 5.13).
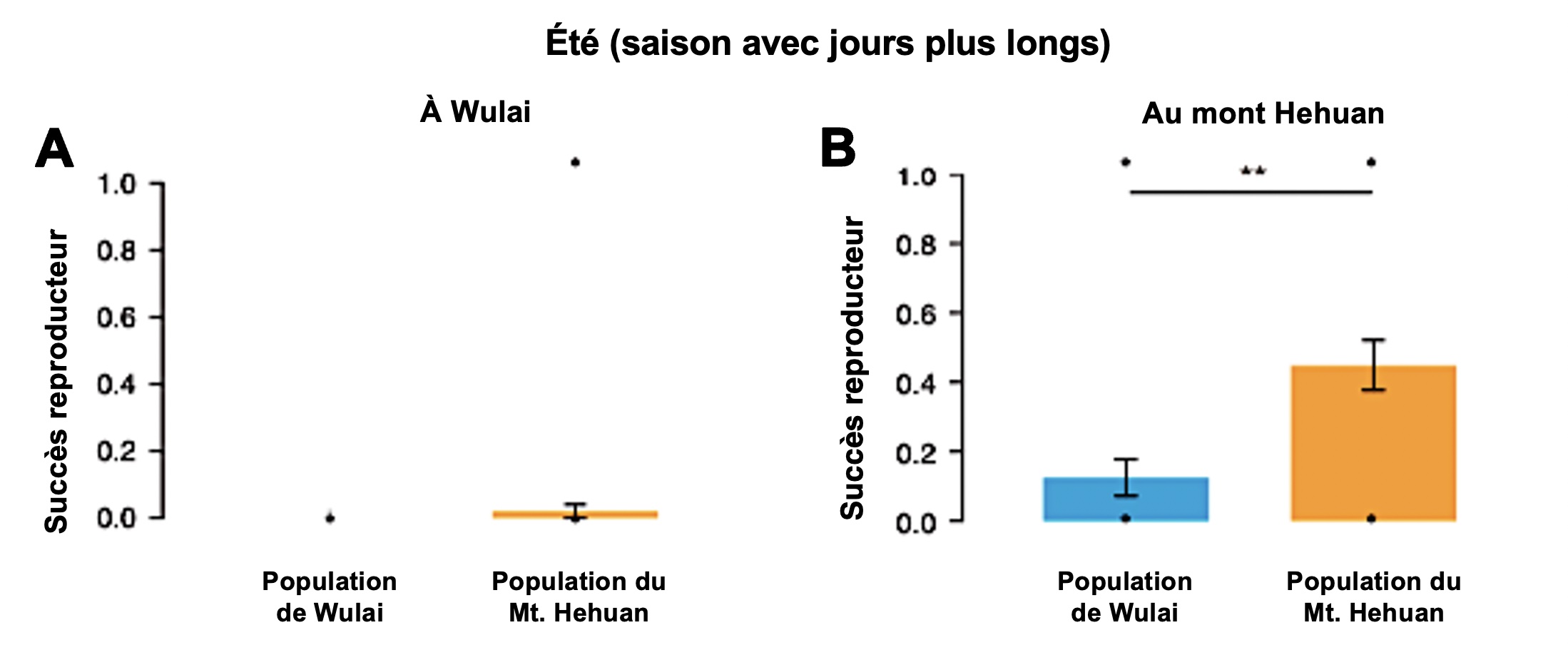
Cette expérience de manipulation de l’environnement par transplantation réciproque a permis de déterminer qu’effectivement le fitness est affecté si on change l’individu d’environnement. Les individus ont d’abord été étudiés durant l’été, avec une longue photopériode. Les individus originaires du Mont Hehuan ont eu un meilleur succès reproducteur que ceux de Wulai lorsqu’ils étaient dans leur territoire d’origine (Figure 5.14).
En contrepartie, lorsque l’expérience avait lieu en hiver, les individus originaires de Wulai avaient un meilleur succès reproducteur que les individus originaires du Mt Hehuan lorsque élevés au Mont Hehuan (figure 5.15).
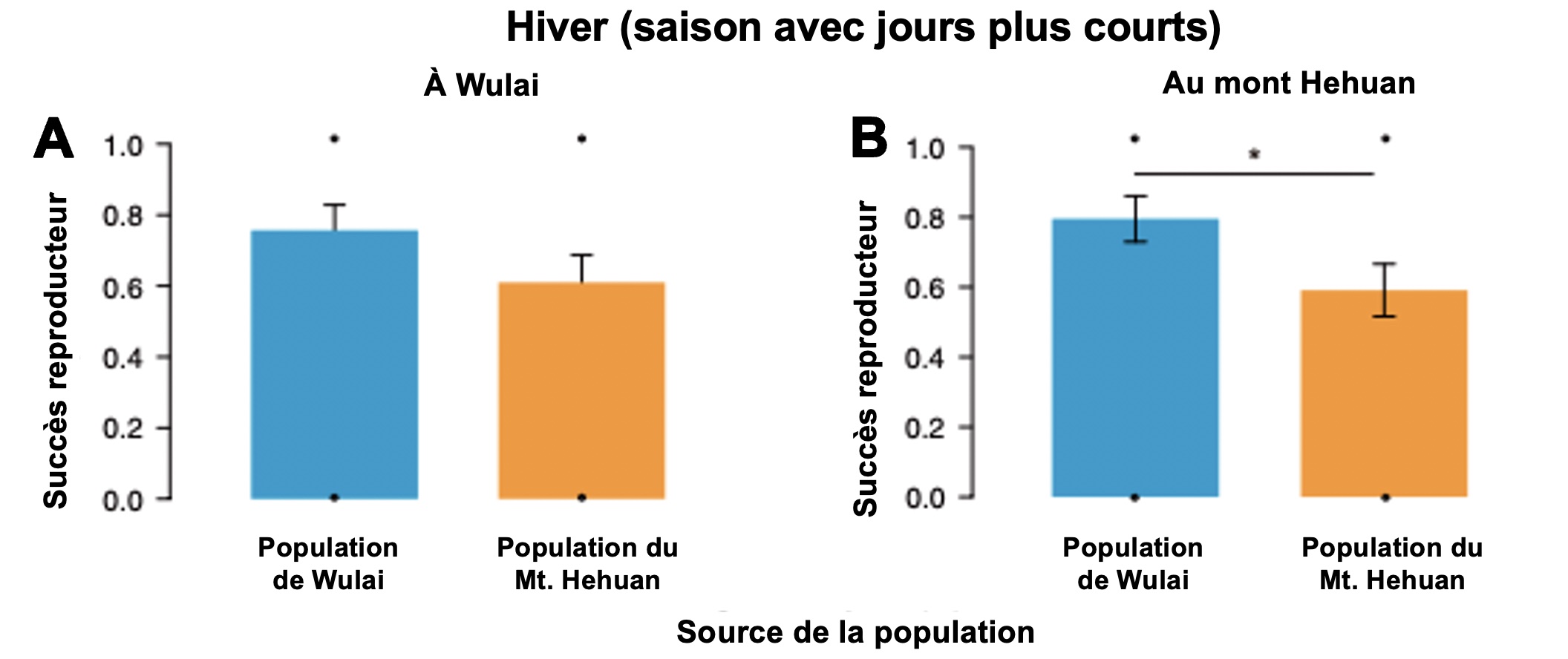
En résumé, les individus du Mont Hehuan se reproduisent sous photopériode courte et longue en laboratoire et ont le meilleur succès reproducteur dans l’étude de transplantation réciproque en photopériode longue (l’été). Les individus de Wulai se reproduisent surtout sous photopériode courte en laboratoire et ont le meilleur succès reproducteur dans l’étude de transplantation réciproque en photopériode courte (l’hiver). Mis ensemble, ces résultats suggèrent que l’activation de la reproduction selon la photopériode est une adaptation locale dans chaque population, car la valeur du trait augmente le fitness par rapport aux autres valeurs du trait dans cet environnement.
Résumé
Une adaptation est un phénotype qui améliore la survie ou la reproduction de l’organisme, relativement aux valeurs alternatives de ce trait, dans un environnement donné. On peut déterminer qu’un trait est une adaptation de trois façons principales : à partir d’observations en nature, en étudiant l’histoire évolutive d’un trait afin de faire ressortir de l’évolution convergente ou divergente, ou en utilisant des méthodes expérimentales où on manipule soit l’environnement, soit le phénotype. Il est aussi possible d’étudier les adaptations à l’aide d’une expérience dévolution expérimentale. Les traits physiologiques qui sont possiblement des adaptations qui permettent de faire face aux défis de l’environnement sont le centre d’intérêt en écophysiologie évolutive. Il est essentiel de vérifier que le trait d’intérêt est effectivement une adaptation.
Activité de révision
Exercices
Révision sur l’étude des adaptations
Bibliographie
- Futuyma, D.J. (2005). Evolution. Sinauer Associates, 3 e édition. 603p.
- Hampshire, G. (2021). Phalène à bouleau [CC-BY]. https://www.flickr.com/photos/gails_pictures/51310547557/in/photolist-2mb8Ya4-2mRoseq-2m7eiZA-2jWQH9P-2jiYixV-2gjE4Lk-2j4gS4u-25pDvvA-2jLh9kS-K82inN-PQ4S9W-2kLX1tH-REUZP5-H9Wnar-2fACUvS-GRJ9DE-2nou8Y3-2j4m3nr-HsXnzU-2nr4Avs-2npESEt-VYxKet-2gaBtCR-2mqXJQi-2jmLGwr-2gz5LXh-2a1eYDU-26N6Rpg-obRSi7-QrfCEu-2gb7UGJ-28Xhh8b-TXwaCo-VgahM5-WwN4aX-2j94ARs-2mwRVao-2mayYuV-HBh55t-2j95Wfr-2nzyG5r-2mx3fr8-28h9vxh-2kYb6Cg-2ntEkx9-2jkN7uT-2m4vej7-2mw5HAb-2njrd8t-2nupVXC
- Sale, B. (2019). Phalène à bouleau [CC-BY]. https://www.flickr.com/photos/33398884@N03/48260144021/in/photolist-2gwzRsD-nA2PsW-2mfy99j-FLRhSy-oq68nS-DMQnLT-oycRTJ-dvzXzD-rAYPPw-m5JxWv-3emrZ-2jmhMrP-ANvSG-PfAZKr-DoAxwW-2jhvZtS-x3XwXy-2kFrewY-DV3wmw-9Z6QMN-m5HVt2-UvUMiA-5W35Zs-2nuePWv-2j5GnSc-284u48o-VtoL6P-2jS5nQ5-24cttSZ-Kc6nbV-2jrxwnS-2gLhFTt-a1gWmF-2jHJSMS-2mgsE35-yQinCY-VtsDbk-4eFUNb-8arTKb-ob8ZzK-4qXdb8-9Qf1wE-WgXFfm-2j9ybVF-W5spY4-VwxJzZ-nPq4At-9BK36F-HxhATx-f9CA45.
- Ketlewel, H.B.D. (1958). A frequency map of Biston betularia and its two melanics, f .carbonaria and f. insularia (1952-56), comprising more than 20,0 records from 83 centres in Britain. [domaine public]. http://faculty.jsd.claremont.edu/dmcfarlane/bio145mcfarlane/PDFs/Kettlewell%201958%20hdy19584a.pdf
- Khan Academy (2022). Sélection naturelle de la population des souris au fil du temps [CC-BY-NC-SA]. https://fr.khanacademy.org/science/biologie-a-l-ecole/x5047ff3843d876a6:bio-4e-annee-sciences-generales/x5047ff3843d876a6:bio-4-2h-l-evolution-et-la-selection-naturelle/a/hs-evolution-and-natural-selection-review
- Schumacher, E.L. et B.A. Carlson (2022). Convergent mosaic brain evolution is associated with the evolution of novel electrosensory systems in teleost fishes [CC-BY]. eLife, 11. https://doi.org/10.7554/eLife.74159
- Romanes, J. (1892). Diagram of homologous bones and analogous flying adaptions in the forelimbs of three groups of flying vertebrates both recent and fossilized: pterosaur (Pterosauria), bat (Chiroptera), bird (Aves) [domaine public]. https://commons.wikimedia.org/wiki/File:Homology.jpg
- Pauling, J. (2022). Red fox [License Pixabay]. https://pixabay.com/photos/fox-red-fox-vulpine-wildlife-fauna-7017260/
- Picard, D. (2016). Arctic fox [License Pixabay]. https://pixabay.com/photos/arctic-fox-animal-mammal-wild-1694817/
- Cifer 88 (2022). Desert fox [License Pixabay]. https://pixabay.com/photos/desert-fox-animal-wildlife-5736089/
- Andersson, M. (1982). Female choice selects for extreme tail length in a widowbird [CC-BY]. Nature, 299, 818-820. https://doi.org/10.1038/299818a0
- Chen, Y., A. Vogel, C. Wagg, T. Xu, M. Iturrate-Garcia, M. Scherer-Lorenzen, A. Weigelt, N. Eisenhauer et B. Schmid (2022). Drought-exposure history increases complementarity between plant species in response to a subsequent drought [CC-BY]. Nature Communications, 13(1), 3217. https://doi.org/10.1038/s41467-022-30954-9
- Fidenci, P. (2009). Phrynosoma platyrhinos [CC-BY-SA]. https://commons.wikimedia.org/wiki/File:Phrynosoma_platyrhinos01.jpg
- Domínguez-Guerrero, S.F., F.R. Méndez-de la Cruz, N.L. Manríquez-Morán, M.E. Olson, P. Galina-Tessaro, D.M. Arenas-Moreno, A. Bautista- del Moral, A. Benítez-Villaseñor, H. Gadsden, R.A. Lara-Reséndiz, C.A. Maciel-Mata, F.J. Muñoz-Nolasco, R. Santos-Bibiano, J.H. Valdez-Villavicencio, G.A. Woolrich-Piña et M.M. Muñoz (2022). Exceptional parallelisms characterize the evolutionary transition to live birth in phrynosomatid lizards [CC-BY]. Nature Communications, 13(1), 2881. https://doi.org/10.1038/s41467-022-30535-w
- Varathan, L. et C. Perl (2019). Nicrophorus nepalensis specimen [CC-BY]. https://commons.wikimedia.org/wiki/File:Nicrophorus_nepalensis_specimen.jpg
- Tsai, H.-Y., D.R. Rubenstein, Y.-M. Fan, T.-N. Yuan, B.-F. Chen, Y. Tang, I.C. Chen et S.-F. Shen (2020). Locally-adapted reproductive photoperiodism determines population vulnerability to climate change in burying beetles [CC-BY]. Nature Communications, 11(1), 1398. https://doi.org/10.1038/s41467-020-15208-w

{kind=link}
{kind=link}
{kind=link}