
3. Les quatre forces évolutives
Introduction au sujet
Dans ce chapitre, nous continuerons notre introduction à l’écophysiologie évolutive en révisant des notions sur l’évolution et les quatre forces évolutives. Nous allons aussi aborder des notions plus spécifiques qui concernent l’évolution par sélection naturelle.
Objectifs spécifiques
À la fin de ce chapitre, vous serez en mesure de :
- Définir ce qu’est une cause ultime
- Définir le concept d’évolution
- Définir les 4 forces évolutives
- Contraster l’évolution neutre et la sélection naturelle
- Définir la sélection naturelle
- Représenter graphiquement une fonction de fitness
- Décrire comment on estime un coefficient de sélection et ce qu’il représente
- Définir le concept d’agent de sélection
- Définir le concept de cible de sélection
- Définir le concept de performance
- Expliquer le lien trait-performance-fitness
Notions clés
1. Les causes ultimes
Le but de l’écophysiologie évolutive est d’étudier l’évolution du fonctionnement des systèmes physiologiques dans différents environnements. Afin d’illustrer plusieurs principes en évolution et en physiologie, nous nous concentrerons sur des adaptations physiologiques qui permettent d’utiliser des habitats avec des conditions environnementales extrêmes et sur des cas de populations ou espèces qui ont divergé drastiquement de leur forme ancestrale au niveau physiologique lorsqu’ils ont envahi de nouveaux habitats. Par exemple, nous verrons comment certains humains sont adaptés à faire face à l’hypoxie en altitude, l’évolution de la vision des couleurs chez les vertébrés, ainsi que de la résistance au froid chez les plantes et les animaux. Ces adaptations impliquent différents systèmes physiologiques, tels que les systèmes circulatoire, digestif, et sensoriel, ainsi que des changements aux niveaux moléculaire et cellulaire. L’étude des causes proximales, qui vise à déterminer comment ces systèmes physiologiques sont modifiés dans leur fonctionnement est un aspect central de la physiologie. Cette question du « comment » formera une grande part de ce texte. En écophysiologie évolutive, l’étude de pourquoi ces traits sont apparus durant l’évolution est aussi d’intérêt. Lorsqu’on se demande « Quelle est la cause évolutive de l’existence de ce trait? », on parle des causes ultimes. Par exemple, on cherchera à déterminer quel est l’agent de sélection, quelle est la cible de la sélection et quel est le lien entre le phénotype d’un individu et son fitness. On peut aussi s’intéresser aux contraintes génétiques (pléiotropie, liaison génétique), phylogénétiques et physiologiques qui posent des limites aux valeurs que peut prendre un trait.
Rien en biologie n’a de sens sauf à la lumière de l’évolution. Vous avez peut-être déjà entendu cette phrase qui est une citation d’un article de Theodosius Dobzhansky, un chercheur en biologie évolutive (figure 3.1). Son article s’adressait aux enseignant.es de biologie à l’école secondaire aux États-Unis [1]. Il voulait les convaincre que pour enseigner la biologie, il faut enseigner l’évolution. On peut réinterpréter le sens de cette phrase et se dire que pour comprendre l’écophysiologie, il faut toujours garder en tête l’étude des causes ultimes en tentant de répondre à la question « pourquoi » ce trait est comme il est, tout en étudiant les causes proximales, c’est-à-dire « comment ».

2. L’évolution
L’évolution se définit comme « un changement dans les fréquences alléliques d’une population à travers les générations » (figure 3.2).
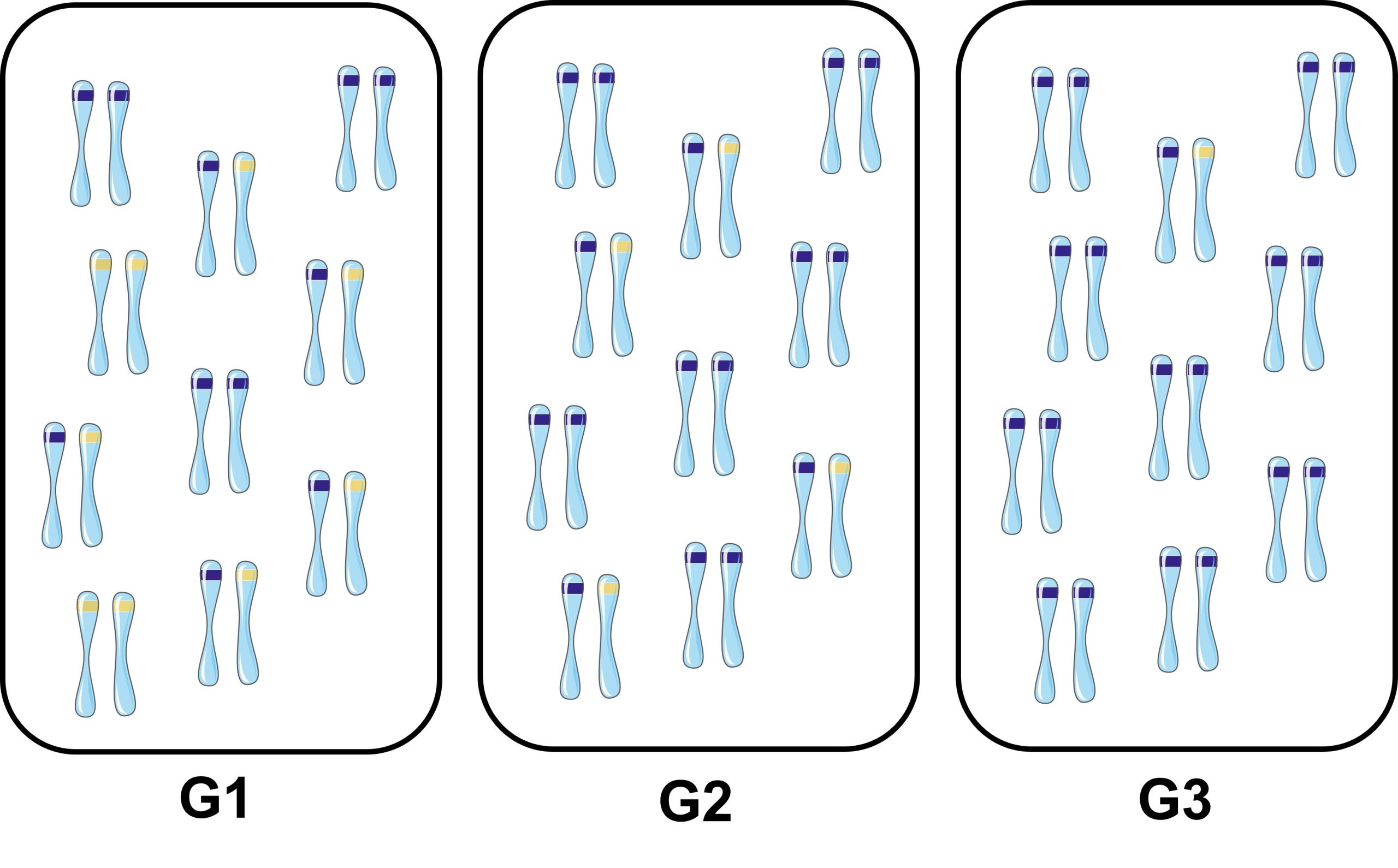
3. Les forces évolutives
La section sur les goulots d’étranglement et les effets fondateurs est adapté de [4].
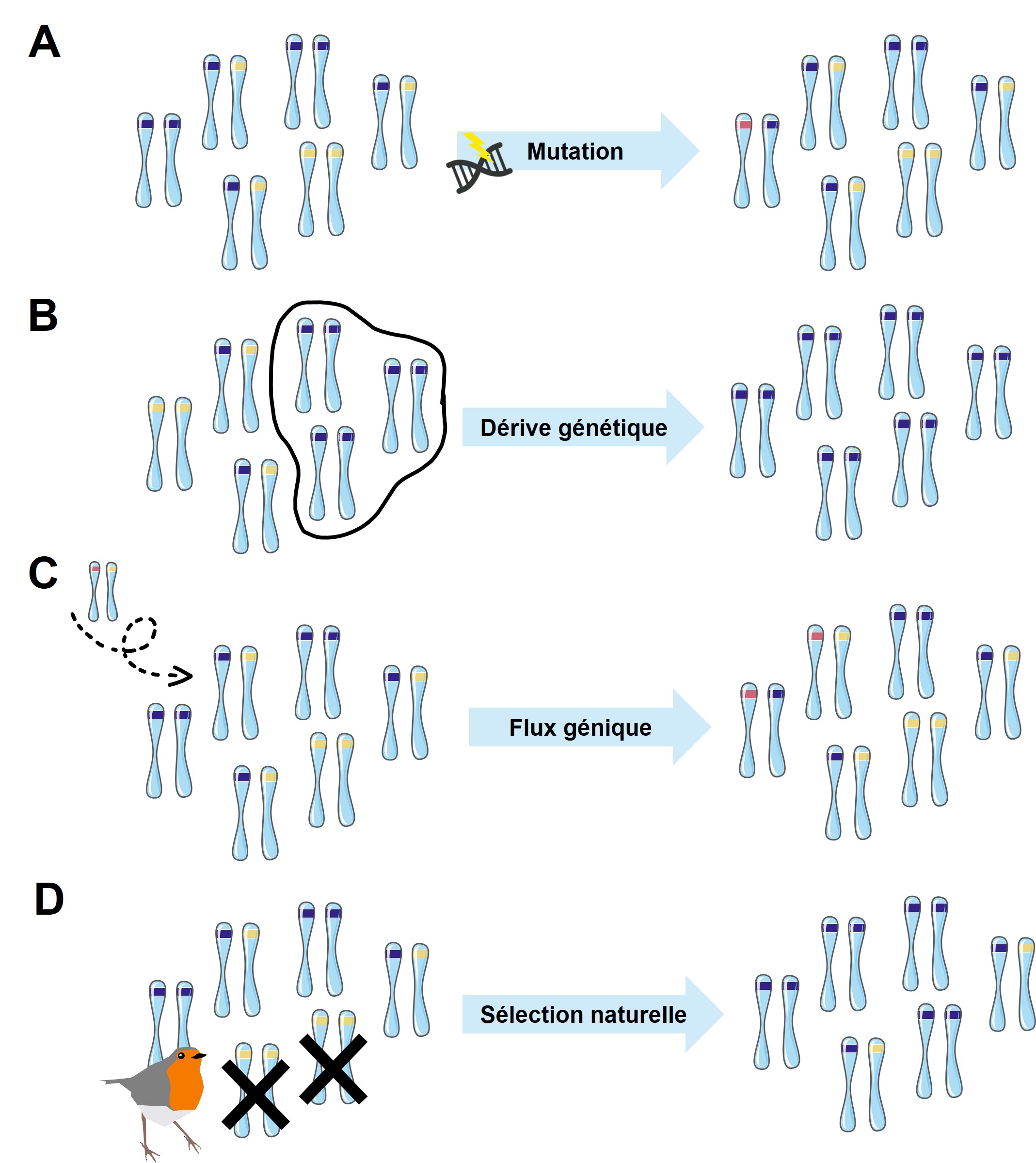
Les forces évolutives qui peuvent causer ces changements de fréquences alléliques entre les générations dans une population sont les mutations, la dérive génétique, le flux génique (qu’on désigne aussi par « migration ») et la sélection naturelle (figure 3.3).
La mutation cause un changement de fréquences alléliques en créant de nouveaux allèles par des changements aléatoires de la séquence de nucléotides (figure 3.3A). Si les mutations sont transmises à la prochaine génération par les gamètes, il y a alors un changement de fréquences alléliques dans cette population.
La dérive génétique résulte d’un échantillonnage des allèles présents dans une population qui ne représente pas parfaitement les fréquences dans celle-ci (figure 3.3B). Cela se produit puisque certains individus se reproduisent et d’autres non et que ceux qui ont des descendants ne représentent pas nécessairement les fréquences alléliques exactes de la population. La dérive se produit dans toutes les populations et c’est l’intensité de la dérive qui varie, principalement selon la taille de la population. Cet échantillonnage inégal de certains allèles versus d’autres entre deux générations modifie les fréquences alléliques. Il y a deux cas particuliers de dérive génétique : les goulots d’étranglement et les effets fondateurs. Un goulot d’étranglement démographique se produit lorsque le nombre d’individus d’une population chute de manière spectaculaire en raison d’un événement aléatoire. Les exemples les plus évidents et les plus connus sont les catastrophes naturelles. Les tsunamis et les ouragans qui dévastent les populations insulaires et côtières, ainsi que les incendies de forêt et les inondations fluviales qui anéantissent les populations d’autres régions, sont des exemples familiers. Lorsqu’une grande partie d’une population est anéantie de manière aléatoire, les fréquences alléliques (c’est-à-dire les pourcentages de chaque allèle) dans la petite population de survivants sont souvent très différentes des fréquences dans la population d’avant la catastrophe, ou « parent ». Les goulots d’étranglement sont donc un cas particulier de dérive génétique. Les effets fondateurs se produisent quant à eux lorsque les membres d’une population quittent le groupe principal et forment une nouvelle population qui ne se croise plus avec les autres membres du groupe d’origine. Comme les survivants d’un goulot d’étranglement de population, la population nouvellement fondée a souvent des fréquences d’allèles différentes de celles du groupe d’origine. Des allèles qui étaient peut-être relativement rares dans la population parente peuvent finir par être très courants en raison de l’effet fondateur. De même, des traits récessifs qui étaient rarement observés dans la population parente peuvent être fréquemment observés chez les descendants de la population dérivée. Ces cas de dérive génétique sont très importants, entre autres lorsqu’on étudie des populations en déclin que l’on tente de conserver et dont l’habitat a été morcelé.
Le flux génique modifie les fréquences alléliques quand un individu avec un certain génotype migre dans une autre population (figure 3.3C). S’il se reproduit, ses allèles contribuent au pool de génotypes de la population d’accueil et modifient les fréquences alléliques dans la population à la nouvelle génération.
La sélection naturelle affecte les fréquences alléliques d’une population si la différence de phénotype entre les individus qui est associée à une différence de fitness est causée par une différence génétique entre les individus (figure 3.3D). Dans ce cas, les individus dont le phénotype est associé à un meilleur fitness transmettent leur génotype à la génération suivante plus fréquemment et les fréquences alléliques changent. La sélection naturelle est la seule force évolutive qui peut mener à l’évolution d’une adaptation. Les trois autres processus peuvent accélérer ou ralentir l’effet de la sélection, mais ne peuvent pas causer une adaptation. Nous discuterons en détail de sélection naturelle dans les sections et chapitres suivants.
Aubin-Horth, N. (2024). Écophysiologie évolutive: les quatre forces évolutives. [Vidéo]. Youtube. CC-BY. https://youtu.be/s_JnDYJsW5U?si=q_G08a8rqrVRPmB3
Vous trouverez la transcription de l’audio de cette capsule en cliquant sur ce lien: Transcription en format word
3.1. L’évolution neutre
Les quatre forces mentionnées peuvent entrainer un changement de fréquences alléliques entre deux générations, et donc une évolution. Ainsi, il est important de se rappeler que, durant l’évolution, les fréquences alléliques et génotypiques changent, mais que ce n’est pas nécessairement à cause de la sélection. Ça peut être à cause des trois autres forces évolutives : la dérive génique, la mutation et l’introduction de migrants (c.a.d le flux génique). Lorsque l’une de ces trois forces crée des changements de fréquences alléliques entre les générations, on parle d’évolution neutre. Ainsi, le changement de fréquences alléliques entre les générations peut être le résultat de la sélection naturelle, mais pas uniquement, ce qui est un aspect à ne pas négliger.
3.2. La sélection naturelle
La sélection naturelle se définit comme une corrélation entre la valeur d’un trait et le fitness. Si on décortique cette définition, on voit d’abord que la valeur d’un trait est importante. Ceci suggère qu’il y a plusieurs valeurs pour ce trait, que les individus n’ont pas tous le même phénotype. Autrement dit, il y a de la variation phénotypique. La deuxième composante de la définition est le fitness. Le fitness est défini comme la capacité d’un individu à survivre et à se reproduire. On parle souvent de succès reproducteur à vie de cet individu, mais ce n’est pas facile à obtenir comme donnée, car il faut en effet avoir la capacité de suivre l’individu pendant toute sa vie. Souvent les biologistes utilisent une variable qui « estime » le fitness, comme le nombre de rejetons dans une portée donnée ou la croissance. Lorsqu’on met ensemble les deux composantes de la définition, on voit que la sélection naturelle, c’est simplement une façon de représenter que certains individus qui ont un phénotype particulier ont un meilleur fitness que les individus qui ont une autre valeur pour ce trait (figure 3.4).

3.3. La fonction de fitness
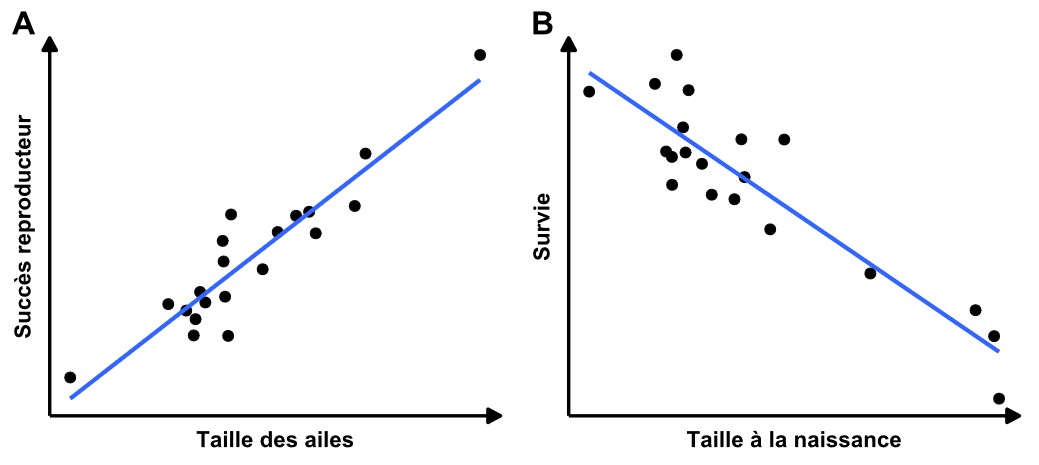
Maintenant que nous connaissons les deux variables qui définissent la sélection naturelle, nous pouvons représenter graphiquement la relation entre les deux. Ce graphique, qui représente la définition de la sélection naturelle, illustre la corrélation entre la valeur d’un trait et le fitness, ce qui permet de voir si ces deux variables sont associées. Par exemple, plus une variable augmente, plus l’autre augmente, ou plus une variable augmente, plus l’autre diminue (figure 3.5). On peut interpréter ces deux graphiques et déterminer ce qu’ils suggèrent d’un point de vue biologique. Dans la figure 3.5A, on observe un phénotype (la taille des ailes) sur l’axe des x qui varie entre les individus et, sur l’axe des y, on a une mesure du fitness, ici le succès reproducteur. Il y a une corrélation positive entre la valeur du phénotype et la variation du fitness : plus un individu a de grandes ailes, meilleur est son fitness. Ce que l’on voit dans ce graphique, c’est la présence de sélection naturelle et plus spécifiquement un exemple de sélection directionnelle. Dans la figure 3.5B, il y a une corrélation négative entre la valeur du phénotype (la taille à la naissance) et la valeur du fitness représenté par la survie : les individus les plus petits à la naissance ont le meilleur fitness.
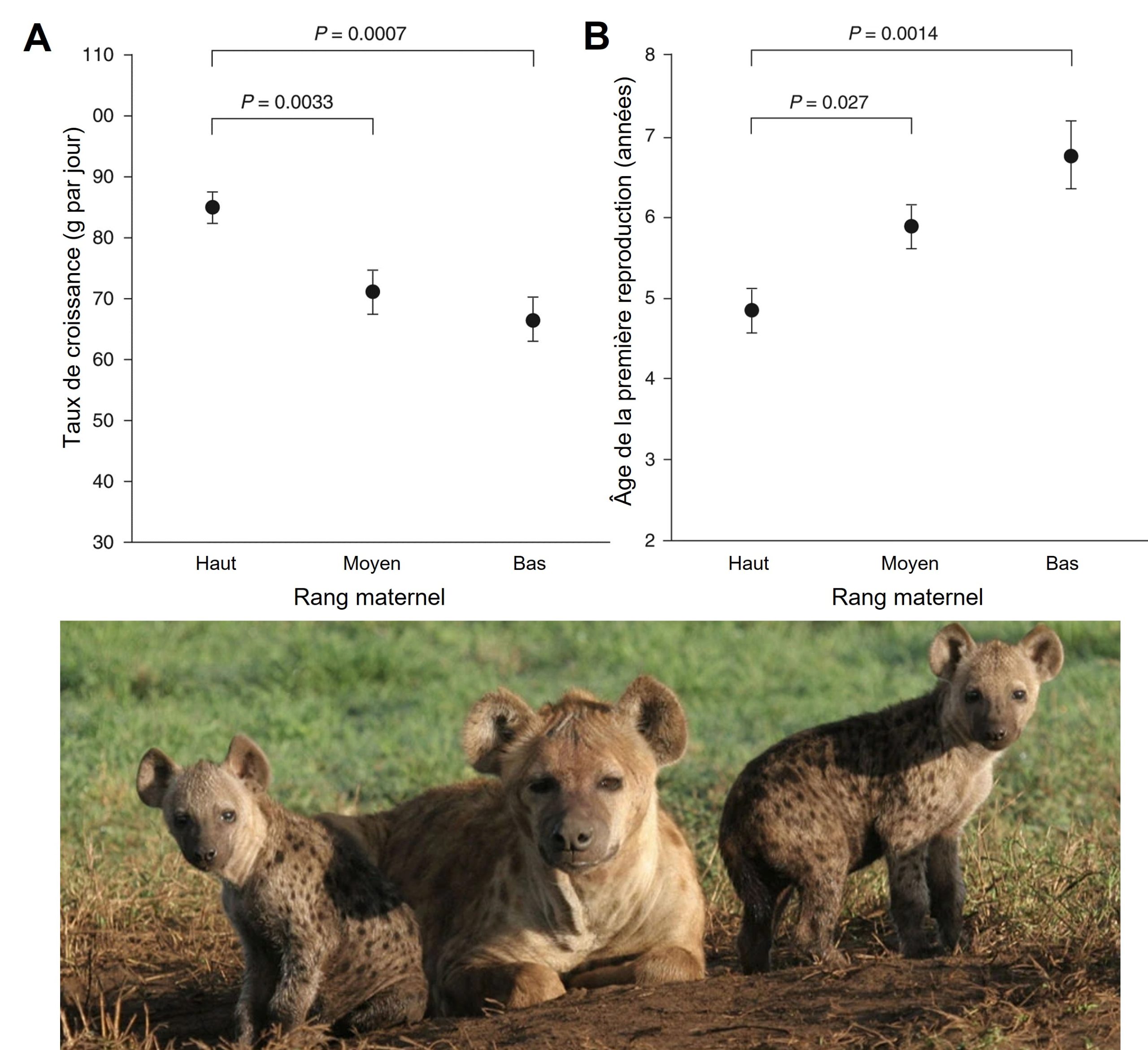
Dans cette figure nous présentons des exemples idéalisés, mais ce type de relation est aussi présent dans la littérature scientifique (figure 3.6).
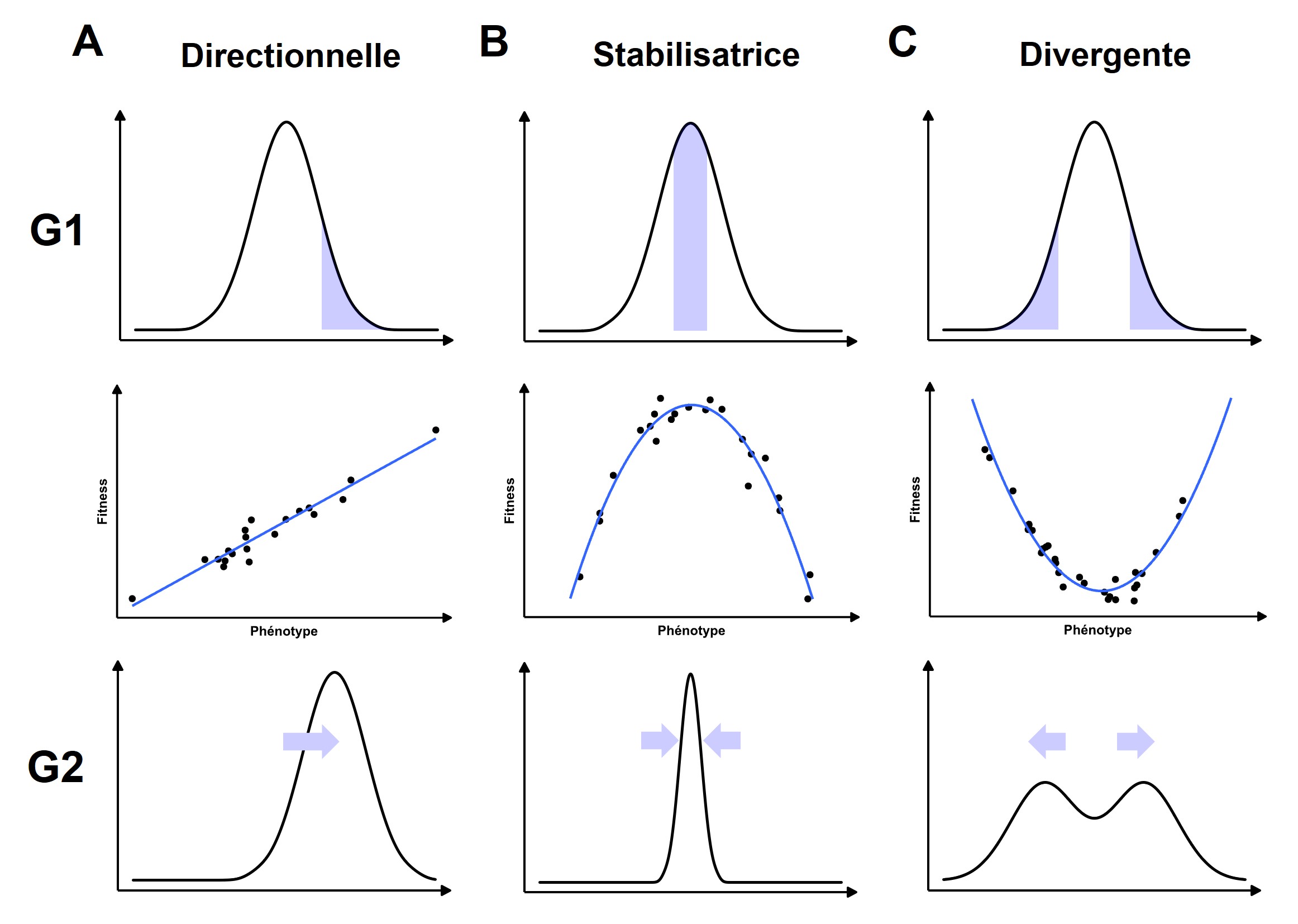
La relation entre le phénotype et la mesure du fitness s’appelle la fonction de fitness. Lorsqu’on étudie une situation de sélection directionnelle, où il y a une relation linéaire entre la valeur du trait et le fitness, la pente de cette fonction nous permet de mesurer le coefficient de sélection (S). La pente représente la force de la sélection, son intensité (figure 3.7A). Nous nous servirons du coefficient de sélection calculé pour la sélection directionnelle lorsque nous estimerons la réponse à la sélection. Prenons des exemples où le trait étudié a une haute héritabilité, ce qui veut dire qu’on peut prédire la valeur du trait pour un rejeton à l’aide de la valeur de ses parents. Nous verrons l’héritabilité en détail plus loin. Dans un cas de sélection directionnelle, seuls les individus avec les valeurs les plus extrêmes vont avoir un fitness élevé dans la génération 1, ce qui déplacera la médiane de la distribution de fréquence vers cette valeur à la génération 2, si le trait a une haute héritabilité (figure 3.7A). Il existe d’autres formes de sélection où la fonction de fitness n’est pas linéaire. Dans le cas de la sélection stabilisatrice, les valeurs extrêmes des phénotypes ont le plus bas fitness à la génération 1 et la relation entre le phénotype et le fitness a la forme d’une parabole (figure 3.7B). Si le trait a une haute héritabilité, la sélection stabilisatrice entraine une diminution de la variance autour de la moyenne à la génération 2 . À l’opposé, on trouve la sélection divergente, où les valeurs extrêmes d’un phénotype sont associées à un meilleur fitness que la valeur médiane et la fonction de fitness a la forme d’une parabole (figure 3.7C). Si le trait a une haute héritabilité, ceci entraine une distribution de fréquence bimodale dans la génération suivante.
Exercices
Vérifiez votre compréhension du concept de fonction de fitness et de coefficient de sélection avec ce quiz formatif.
Le coefficient de sélection
3.4. L’agent de sélection et la cible de la sélection
Les différences de fitness entre les individus qui sont associées à des différences dans leur phénotype sont causées ultimement par des facteurs biotiques et abiotiques de l’environnement. En effet, le fait qu’une certaine valeur de trait soit associée avec un meilleur fitness est dû à une variable de l’environnement qui crée une pression de sélection sur les individus de cette population. Ces facteurs sont les agents de sélection, c’est-à-dire la cause environnementale des différences de fitness entre organismes qui ont des phénotypes différents. Ces agents de sélection peuvent être des prédateurs, des facteurs abiotiques comme la température ou l’apport en eau, ou encore des pathogènes (figure 3.8).

La cible de la sélection est le trait phénotypique sur lequel la sélection agit directement. Autrement dit, la cible de sélection est un trait qui aide l’organisme à faire face à l’agent de sélection, c’est-à-dire, l’environnement. La cible peut donc être tout trait à n’importe quel niveau d’organisation biologique qui va affecter le fitness dans cet environnement (figure 3.9). Attention : on ne peut pas savoir si un trait est effectivement la cible de la sélection sans avoir fait des expériences supplémentaires demandant des manipulations, afin de vérifier qu’effectivement c’est la valeur de ce trait spécifiquement qui affecte le fitness.

3.5. La performance
Lorsqu’on étudie la relation entre le phénotype et le fitness, on a souvent comme prémisse implicite que c’est ce trait qui affecte le fitness, qu’il est la cible de la sélection. Par exemple, on mesure qu’une différence dans la taille et la forme de la mâchoire entre des individus d’une même population de serpent sont associées à une différence de survie. Il est intéressant de disséquer cette relation en deux parties afin de rendre plus explicite le lien entre ces deux variables : 1) le phénotype a un effet sur la performance de l’organisme et 2) la performance de l’organisme a un effet sur le fitness. Dans le cas de notre serpent, qui fût l’exemple présenté originalement pour illustrer ce concept par Steven Arnold dans les années 80, la longueur et la forme des différents os qui forment la mâchoire et la boite crânienne sont un ensemble de traits qui affectent la performance d’ingestion, estimée par la circonférence maximale de la proie qui peut être ingérée. Ce phénotype (la longueur et la forme des os de la mâchoire) varie entre les individus et affecte leur performance, soit la capacité à s’alimenter d’une proie d’une certaine taille en l’avalant entier. Les individus qui ont une meilleure performance sont ceux qui peuvent se nourrir de ces proies d’une certaine taille et qui ont une meilleure survie et une meilleure reproduction. Il y a donc un premier lien phénotype-performance (taille et forme de la mâchoire-performance d’ingestion) et un deuxième lien performance-fitness (taux d’ingestion-survie).
Comme vous pouvez le remarquer, le taux d’ingestion implique la morphologie du serpent (elle-même composée de plusieurs os), ainsi que son comportement et sa physiologie (taille et force de la musculature). En effet, la performance est souvent le résultat de la combinaison de la valeur de plusieurs traits. On étudie souvent le premier lien (phénotype-performance) en laboratoire, bien que de plus en plus d’études se font en nature, alors que le plus souvent le deuxième lien performance-fitness doit être testé en conditions naturelles ou expérimentales très proches de l’environnement naturel afin d’estimer correctement le fitness. La performance est un concept qui est utilisé en écologie (vous avez peut-être déjà entendu parler de « traits fonctionnels »), en évolution, dû à son lien direct avec la sélection naturelle, ainsi qu’en physiologie, où on mesure souvent la performance face à des défis de l’environnement, tel que la température. La performance est en quelque sorte la « connexion » entre les niveaux d’organisation biologique sous-organismiques (le niveau physiologique, lui-même affecté par les niveaux cellulaires et moléculaires), l’organisme entier, l’environnement et le fitness. En physiologie, la performance est souvent mesurée avec des processus biologiques qui ont une composante temporelle : la vitesse de course, le taux de battements cardiaques, la quantité d’oxygène utilisée, la croissance, ou l’activité enzymatique. D’ailleurs, la publication du premier article présentant le cadre conceptuel proposant de relier le trait, la performance et le fitness a été un point tournant dans le développement du champ de l’écophysiologie évolutive dans les années 1980. Étudier la performance nous permet en quelque sorte « d’expliquer » pourquoi on observe une relation entre un phénotype et le fitness.
Cas appliqué
L’évolution neutre
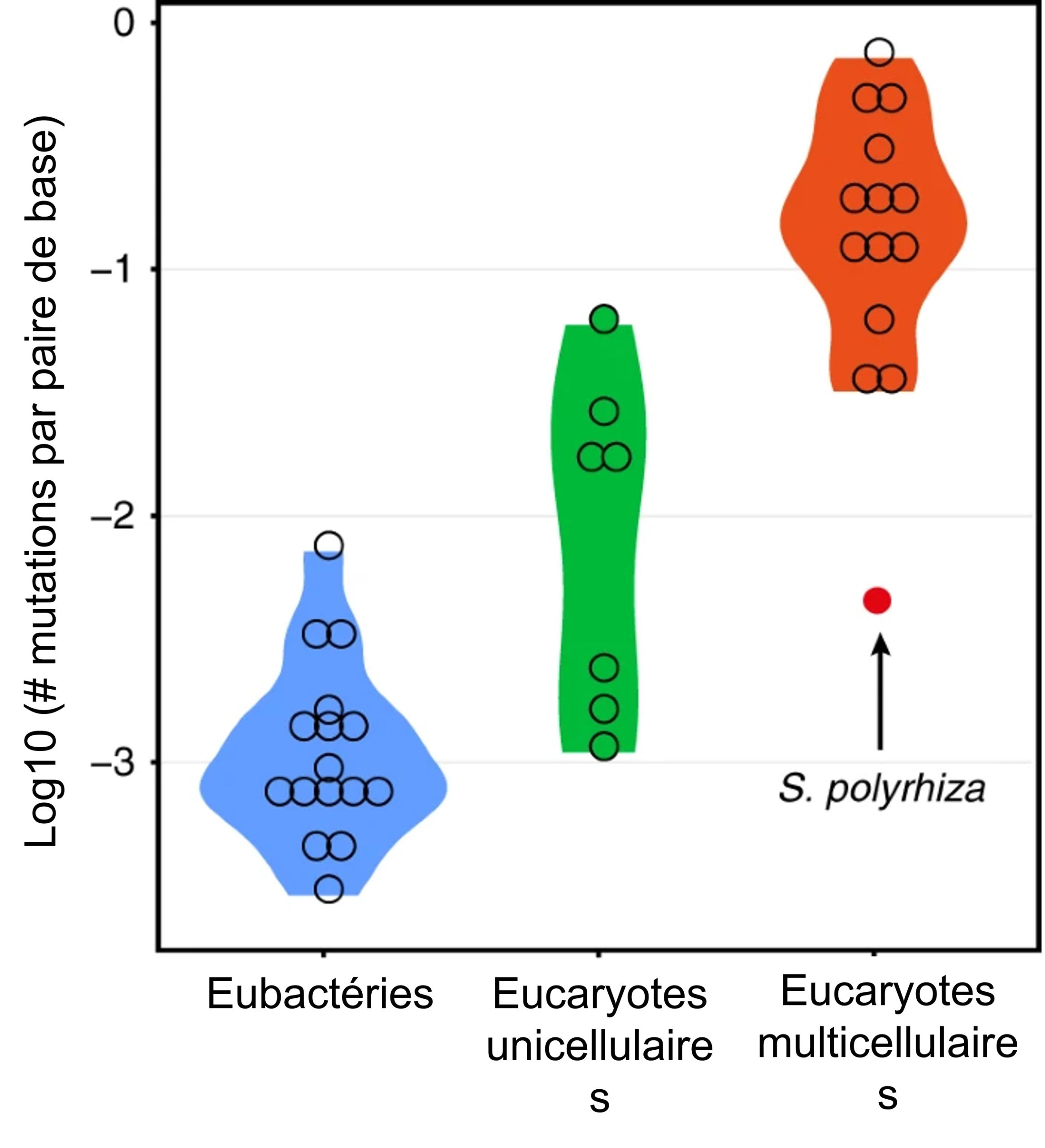
Nous avons vu que les mutations sont une des forces évolutives qui causent de l’évolution neutre. En effet, une mutation va causer un changement de fréquence allélique entre les générations, qui est la définition de l’évolution. Des biologistes ont estimé le taux de mutation d’une espèce de plante, la lentille d’eau (Spirodela polyrrhiza) (figure 3.10) afin de le comparer à celui d’autres espèces. Elles ont mesuré expérimentalement le taux de mutation dans le génome de lentilles d’eau à l’aide d’une expérience d’accumulation de mutations. Elles ont fait ces expériences en recréant des conditions naturelles, par exemple de UV et de température, afin d’avoir un estimé juste du taux de mutation dans cette espèce retrouvée pratiquement partout dans le monde et qui a des tailles de population très importantes.

Les biologistes ont comparé le taux de mutation quantifié pour les régions codantes du génome pour différents groupes : les eubactéries, les eucaryotes unicellulaires et les eucaryotes multicellulaires. Parmi ces derniers, elles ont pu répertorier une grande variation dans le taux de mutation (figure 3.11). Grâce à leur expérience d’accumulation de mutations, elles ont déterminé que la lentille d’eau a un taux de mutation différent des autres espèces incluses dans ce groupe. Le fait que cette espèce soit différente n’est pas tellement important dans cet exemple, c’est plutôt une façon de vous rappeler qu’il y a des mutations constamment dans le génome des individus et que certaines sont transmises à la prochaine génération.
Dérive génique ou sélection?
Des scientifiques ont étudié le chou champêtre (Brassica rapa) (figure 3.12) afin de déterminer si des changements de phénotypes à travers les générations sont le résultat de la dérive génique ou plutôt le résultat de la sélection naturelle lié au type de pollinisateur.

Les scientifiques ont utilisé une approche d’évolution expérimentale. Ils ont créé des lignées de choux et les ont reproduites pendant 9 générations en utilisant comme pollinisateur un bourdon ou une petite mouche, afin de quantifier quels sont les traits qui évoluent avec chaque type de pollinisation (figure 3.13). Ils ont aussi utilisé une lignée témoin pollinisée à la main. Ils ont créé 3 réplicats pour chaque type de pollinisateur, afin de déterminer si les changements de moyenne de phénotype observés sont le résultat de la sélection ou simplement de la dérive génique. Selon eux, si tous les réplicats évoluent dans la même direction, cela supporte l’hypothèse que les changements de phénotypes sont le résultat de la sélection. Si des réplicats du même traitement de pollinisation n’ont pas les mêmes phénotypes après 11 générations, ils déterminent que c’est de l’évolution due à de la dérive génique.

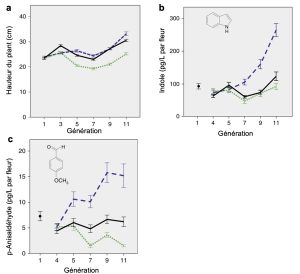
Les biologistes ont observé une divergence phénotypique au fil des générations entre les lignées avec différents types de pollinisateurs, et ce pour différents traits. Après 11 générations, les plantes pollinisées par les bourdons avait une augmentation significative de taille (figure 3.14A) et de quantité d’indole produite (figure 3.14B). L’indole est une molécule qui compose l’odeur de la plante. La quantité de p-anisaldehyde, une autre molécule composant l’odeur de la plante, avait aussi augmentée chez les lignées pollinisées par les bourdons par rapport à la lignée témoin, mais avait diminué significativement chez les plantes pollinisées par la mouche (figure 3.14C). La moitié des plantes pollinisées par la mouche ne produisait plus du tout cette molécule après 11 générations. Tous les réplicats d’un même traitement de pollinisation se ressemblaient plus entre eux et avaient évolué dans la même direction, ce qui fait dire aux scientifiques que les changements observés entre générations sont le résultat de la sélection naturelle et non de la dérive génique.
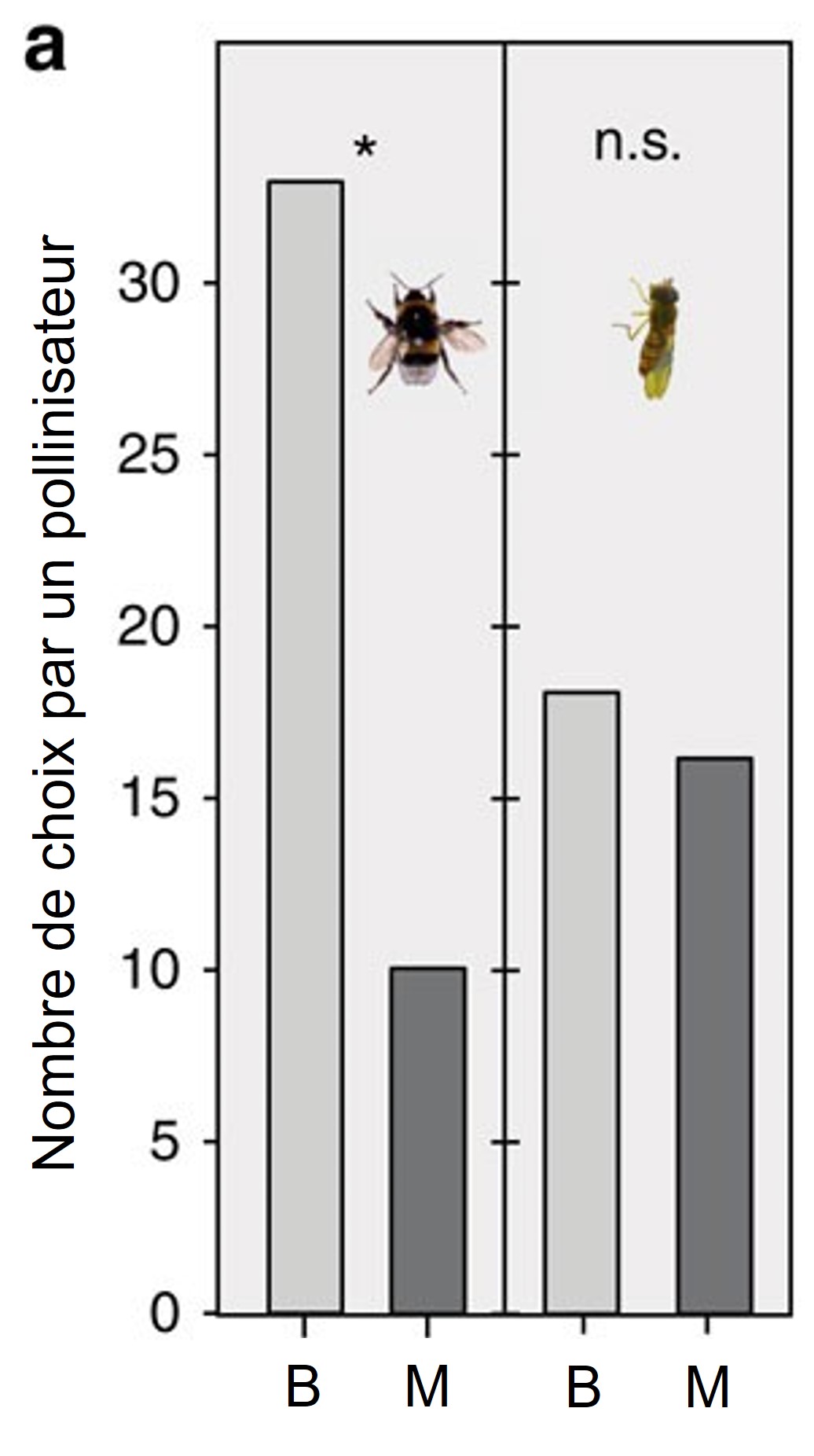
Les biologistes ont calculé des coefficients de sélection significatifs pour plusieurs traits dans chaque traitement de pollinisation. Ils ont aussi noté que certains traits comme la taille de la plante et la quantité de molécules odorantes étaient corrélés avec la quantité de nectar de la plante, ce qui suggère que ces traits étaient un « signal » que le bourdon pouvait utiliser pour choisir une plante lui donnant plus de nectar. Après 11 générations, si on donnait le choix à un bourdon de visiter une plante venant de la lignée qui a évolué avec le bourdon comme pollinisateur (plus grande, plus d’odeur) ou de la lignée qui a évolué avec la mouche, le bourdon naïf choisissait presque toujours la plante qui avait évolué avec les bourdons (figure 3.15). À l’opposé, une mouche naïve n’avait aucune préférence pour une lignée ou l’autre. Les scientifiques concluent que les pollinisateurs peuvent être des agents de sélection importants en nature et que plusieurs traits des plantes peuvent être la cible de la sélection. Ils proposent que si certains pollinisateurs disparaissent, ceci pourrait affecter l’évolution des plantes de façon marquée.
Résumé
Étudier les causes ultimes veut dire déterminer la cause évolutive de l’existence d’un trait, comme l’agent de sélection, la cible de la sélection et le lien entre ce trait et le fitness de l’individu. L’évolution consiste en des changements de fréquences alléliques entre les générations dans une population. Il existe quatre forces évolutives qui peuvent mener à des changements de fréquence allélique entre les générations dans une population : la mutation, la dérive génétique, le flux génique et la sélection naturelle. Les trois premières forces résultent en évolution neutre et seule la sélection naturelle peut résulter en une adaptation. La sélection naturelle est quantifiée comme une corrélation entre la valeur d’un trait d’un individu et le fitness de celui-ci. La représentation graphique de cette relation s’appelle la fonction de fitness. Lorsqu’on étudie la sélection naturelle, on veut déterminer l’agent de sélection, soit la variable environnementale qui cause les différences de fitness entre les phénotypes ainsi que la cible de la sélection, le trait dont la fonction affecte directement le fitness. Finalement, le lien entre le phénotype et le fitness est explicité par l’étude de la performance de l’individu. On peut en effet séparer le lien entre ces deux variables en deux sous-composantes: 1) le phénotype a un effet sur la performance de l’organisme et 2) la performance de l’organisme a un effet sur le fitness. La performance est en quelque sorte la « connexion » entre les niveaux d’organisation biologique sous-organismiques (le niveau physiologique, lui-même affecté par les niveaux cellulaires et moléculaires), l’organisme entier, l’environnement et le fitness.
Activité de révision
Exercices
Révision sur les quatre forces évolutives.
Bibliographie
- Dobzhansky, T. (2013). Nothing in biology makes sense except in the light of evolution. American Biology Teacher, 75(2), 87-91. https://doi.org/10.2307/4444260
- Auteur inconnu (1943). Dobzhansky no Brasil em 1943 [domaine public]. https://commons.wikimedia.org/wiki/File:Dobzhansky_no_Brasil_em_1943.jpg
- Servier. Meiosis [CC-BY]. https://smart.servier.com/smart_image/meiosis/
- Shook, B. Nelson, K. Aguilera K. et L. Braff. (2019). Explorations. CC-BY. https://pressbooks-dev.oer.hawaii.edu/explorationsbioanth/chapter/__unknown__-3/
- Servier. Meiosis [CC-BY]. https://smart.servier.com/smart_image/meiosis/
- Electronic_Frontier_Foundation (2018). DNA-icon [CC-BY]. https://www.flickr.com/photos/59202267@N08/40124398361
- Auteur inconnu. Anger lightning bolt [CC0]. https://svgsilh.com/image/305229.html
- Auteur inconnu (2020). Robin bird grey and orange [domaine public]. https://freesvg.org/robin-bird-grey-and-orange
- Tooony (2007). Principe_variation [CC-BY-SA]. https://commons.wikimedia.org/wiki/File:Principe_variation.svg
- Tooony (2007). Template:Graphical rapresentation of natural selection [CC-BY-SA]. https://commons.wikimedia.org/wiki/File:Selection.svg
- Tooony (2007). Principe d’hérédité [CC-BY-SA]. https://commons.wikimedia.org/wiki/File:H%C3%A9r%C3%A9dit%C3%A9.svg
- Höner, O.P., B. Wachter, H. Hofer, K. Wilhelm, D. Thierer, F. Trillmich, T. Burke et M.L. East (2010). The fitness of dispersing spotted hyaena sons is influenced by maternal social status [CC-BY]. Nature Communications, 1(1), 60. https://doi.org/10.1038/ncomms1059
- Christels (2018). Insecte-mouche-invertebre [License Pixabay]. https://pixnio.com/fr/faune-animaux/insectes-et-bugs/fly-photos/insecte-mouche-invertebre-animal-feuille-detail-macro-metamorphose#
- Rob Lee (2006). Fox in snow [CC-BY]. https://www.flickr.com/photos/55761924@N00/102165753
- Alexandre Keledjian (2003). Plante Desert Figuig [CC-BY-NC]. https://www.flickr.com/photos/29334587@N04/2745123896
- CharlesFred (2006). Apple [CC-BY-NC-SA]. https://www.flickr.com/photos/40279823@N00/253443873
- Brisbane City Council (2012). Mangroves [CC-BY]. https://www.flickr.com/photos/50553864@N06/7114626659
- South African Tourism (2009). Giraffe eating a thorn tree, North West Province [CC-BY]. https://www.flickr.com/photos/25779097@N08/6253217436
- David Cook Wildlife Photography (2009). Sparkling Violet-ear (Colibri coruscans) [CC-BY-NC]. https://www.flickr.com/photos/16520061@N08/3794764350
- David Reed (2011). Giant Swallowtail – Papilio cresphontes [CC-BY-NC-SA]. https://www.flickr.com/photos/60382879@N06/5535207769
- Saij, I. (2005). Spirodela polyrrhiza marais poitevin [License Free Art]. https://commons.wikimedia.org/wiki/File:Spirodela_polyrrhiza_marais_poitevin.jpg
- Xu, S., J. Stapley, S. Gablenz, J. Boyer, K.J. Appenroth, K.S. Sree, J. Gershenzon, A. Widmer et M. Huber (2019). Low genetic variation is associated with low mutation rate in the giant duckweed [CC-BY]. Nature Communications, 10(1). https://doi.org/10.1038/s41467-019-09235-5
- TeunSpaans (2004). Brassica rapa plant [CC BY-SA]. https://commons.wikimedia.org/wiki/File:Brassica_rapa_plant.jpg
- Entomart (2006). Bombus terrestris01 [CC BY]. https://commons.wikimedia.org/wiki/File:Bombus_terrestris01.jpg
- Karwath, A. (2005). This image shows a 12 mm large hoverfly called marmelade fly (Episyrphus balteatus) from top [CC BY-SA]. https://commons.wikimedia.org/wiki/File:Episyrphus_balteatus_male_-_top_(aka).jpg
- Gervasi, D.D.L. et F.P. Schiestl (2017). Real-time divergent evolution in plants driven by pollinators [CC-BY]. Nature Communications, 8(1), 14691. https://doi.org/10.1038/ncomms14691

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}