
16. Le système sensoriel
Introduction
Dans ce chapitre, nous aborderons le système sensoriel dont le rôle est de procurer de l’information sur l’environnement. Pour ce faire, il est composé de sens spécifiques et généraux. Nous verrons les différents types de cellules réceptrices, comment on les classe selon leur localisation et leur modalité sensorielle, ainsi que les étapes de la transduction sensorielle. Nous verrons comment différents aspects de l’information sensorielle, soit le type de stimulus, son emplacement, son intensité et sa durée, sont encodés par les systèmes sensoriels et le système nerveux central. Finalement, nous verrons comment l’information est intégrée et résulte en signal aux effecteurs pour par exemple résulter en une réponse motrice.
Objectifs spécifiques
À la fin de ce chapitre, vous serez en mesure de :
- Définir le rôle du système sensoriel
- Expliquer les étapes de la transduction sensorielle
- Contraster la sensation et la perception
- Définir les caractéristiques communes et les rôles des différents types de cellules réceptrices du système sensoriel.
- Comparer les potentiels générateurs et les potentiels de récepteur
- Classer les récepteurs selon leur localisation et leur modalité sensorielle
- Expliquer comment se fait l’encodage des informations sensorielles
- Expliquer le rôle du thalamus dans l’intégration des informations du système sensoriel
- Expliquer le concept de topographie sensorielle
- Décrire les caractéristiques de la carte corticale
- Expliquer le rôle des régions du lobe frontal dans l’activation de la réponse des effecteurs moteurs
Notions clés
- Le système sensoriel
- La transduction sensorielle
- Sensation et perception
- Types de cellules réceptrices : neurone afférent et cellule non neuronale
- Potentiel générateur et potentiel de récepteur
- Classification des récepteurs : localisation et modalité sensorielle
- Encodage et transmission des informations sensorielles
- Intégration dans le système nerveux central et signal aux effecteurs
- Le thalamus et le cortex cérébral
- Topographie sensorielle
- La carte corticale est proportionnelle à la quantité d’information du système sensoriel et est plastique
- La carte corticale varie entre les espèces
- Réponses motrices
Les sections 1 à 5 sont adaptées de [1-3]. Cette section est donc sous licence [CC-BY-SA] afin de maintenir la licence d’un des textes adaptés.
1. Le système sensoriel
Le rôle du système sensoriel est de procurer de l’information sur l’environnement qui entoure l’organisme ou sur l’état de son environnement interne. Si on prend l’exemple des animaux, nous possédons des sens spécifiques et des sens généraux. Les sens spécifiques sont associés à un organe. Chez l’humain, on pense à l’olfaction, la gustation, l’équilibre, la vision et l’ouïe. Les sens généraux quant à eux reposent sur des récepteurs qui ne sont pas associés à un organe spécialisé, mais sont plutôt répartis dans tout le corps dans une variété d’organes. Ces sensations peuvent donc être internes ou externes. Les récepteurs de la peau, des muscles, des parois des vaisseaux sanguins qui détectent la pression et les sensations viscérales sont des exemples de sens généraux. La somatosensation, également appelée somesthésie (« soma » signifie « relié aux cellules du corps »), est considérée comme un sens général. La somatosensation regroupe les sensations associées au toucher et à la position des membres. Ces sensations comprennent la pression, les vibrations, le toucher léger, les chatouillements, les démangeaisons, la température, la douleur, la proprioception (position des os, des articulations et des muscles) et la kinesthésie (mouvement des membres du corps). De nombreux récepteurs somatosensoriels sont situés dans la peau, mais des récepteurs se trouvent également dans les muscles, les tendons, les capsules articulaires et les ligaments.
2. La transduction sensorielle
Bien que les systèmes sensoriels associés à ces sens soient très différents, tous partagent une fonction commune : convertir un stimulus (comme la lumière, ou le son, ou la position du corps) en un signal électrique dans le système nerveux. Ce processus appelé transduction sensorielle est composé de trois étapes : la réception, la transduction et la perception.
La première étape de la sensation est la réception. Il s’agit de l’activation des récepteurs sensoriels par des stimuli tels que des stimuli mécaniques (être plié ou écrasé, par exemple), des produits chimiques ou la température. La deuxième étape est la transduction. La fonction fondamentale du système sensoriel est la traduction d’un signal sensoriel en un signal électrique dans le système nerveux. Cela se produit au niveau du récepteur sensoriel, et le changement de potentiel électrique qui est produit est un potentiel de récepteur. Comment le signal sensoriel, tel que la pression sur la peau ou la pression d’une onde sonore, est-il transformé en un potentiel de récepteur? Prenons l’exemple d’un type de récepteur appelé mécanorécepteur, que l’on retrouve entre autres dans le système auditif. Il s’agit de neurones particuliers qui possèdent des membranes spécialisées répondant à la pression. Les mécanorécepteurs détectent les stimuli dus à la déformation physique de leurs membranes plasmiques. Ils contiennent des canaux ioniques à ouverture mécanique qui s’ouvrent ou se ferment en réponse à la pression, au toucher, à l’étirement et au son, ce qui modifie le potentiel électrique (figure 16.1A). Rappelons que dans le système nerveux, une variation positive du potentiel électrique d’un neurone (aussi appelé potentiel de membrane) dépolarise le neurone et qu’une variation négative entraine une hyperpolarisation. Les potentiels de récepteur sont des potentiels gradués : l’amplitude de ces potentiels gradués varie avec la force du stimulus et/ou l’addition de plusieurs stimuli. Si l’amplitude de la dépolarisation est suffisante (c’est-à-dire si le potentiel de membrane atteint un seuil), le neurone déclenchera un potentiel d’action (figure 16.1B). Dans la plupart des cas, le stimulus sur un récepteur sensoriel entrainera le potentiel membranaire dans une direction positive (une dépolarisation). Il faut noter que certains récepteurs, tels que ceux du système visuel, sont un cas spécial où il y a plutôt hyperpolarisation.
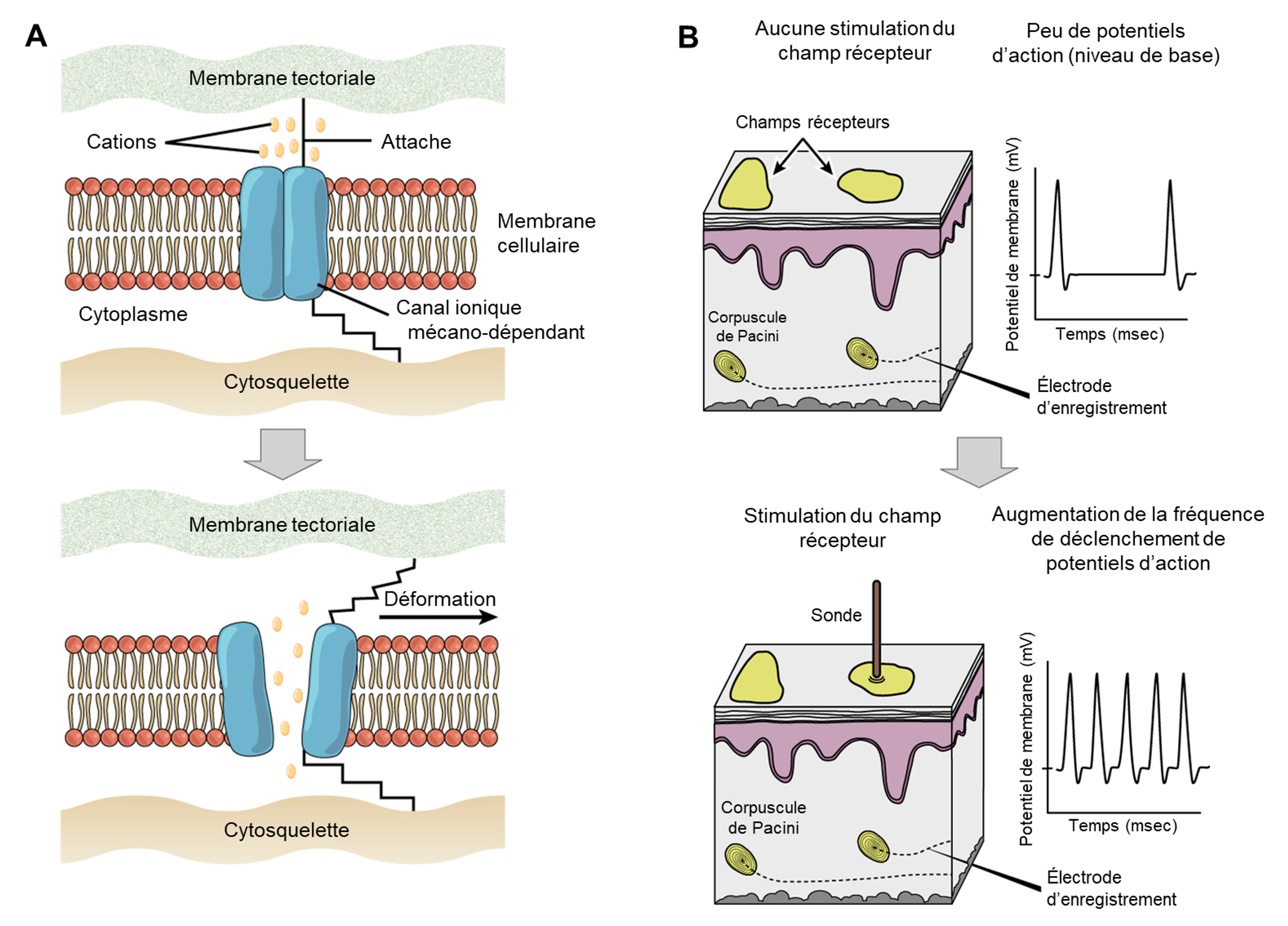
La troisième étape est la perception et se produit une fois que le potentiel d’action est relayé dans le système nerveux central, où il est intégré à d’autres informations sensorielles — et parfois à des fonctions cognitives supérieures — pour devenir une perception consciente de ce stimulus. L’intégration centrale peut alors conduire à une réponse motrice. La perception est l’interprétation d’une sensation par un individu. Bien que la perception repose sur l’activation des récepteurs sensoriels, la perception ne se produit pas au niveau du récepteur sensoriel, mais à des niveaux plus élevés dans le système nerveux, dans le cerveau. Tous les signaux sensoriels, à l’exception de ceux du système olfactif (qui vont directement au cortex cérébral), sont transmis par le système nerveux central et sont acheminés vers le thalamus et vers la région appropriée du cortex. Rappelons que le thalamus est une structure du cerveau antérieur qui sert de centre d’échange et de relais pour les signaux sensoriels (ainsi que moteurs). Lorsque le signal sensoriel sort du thalamus, il est dirigé vers la zone spécifique du cortex (figure 16.2) dédiée au traitement de ce sens particulier.
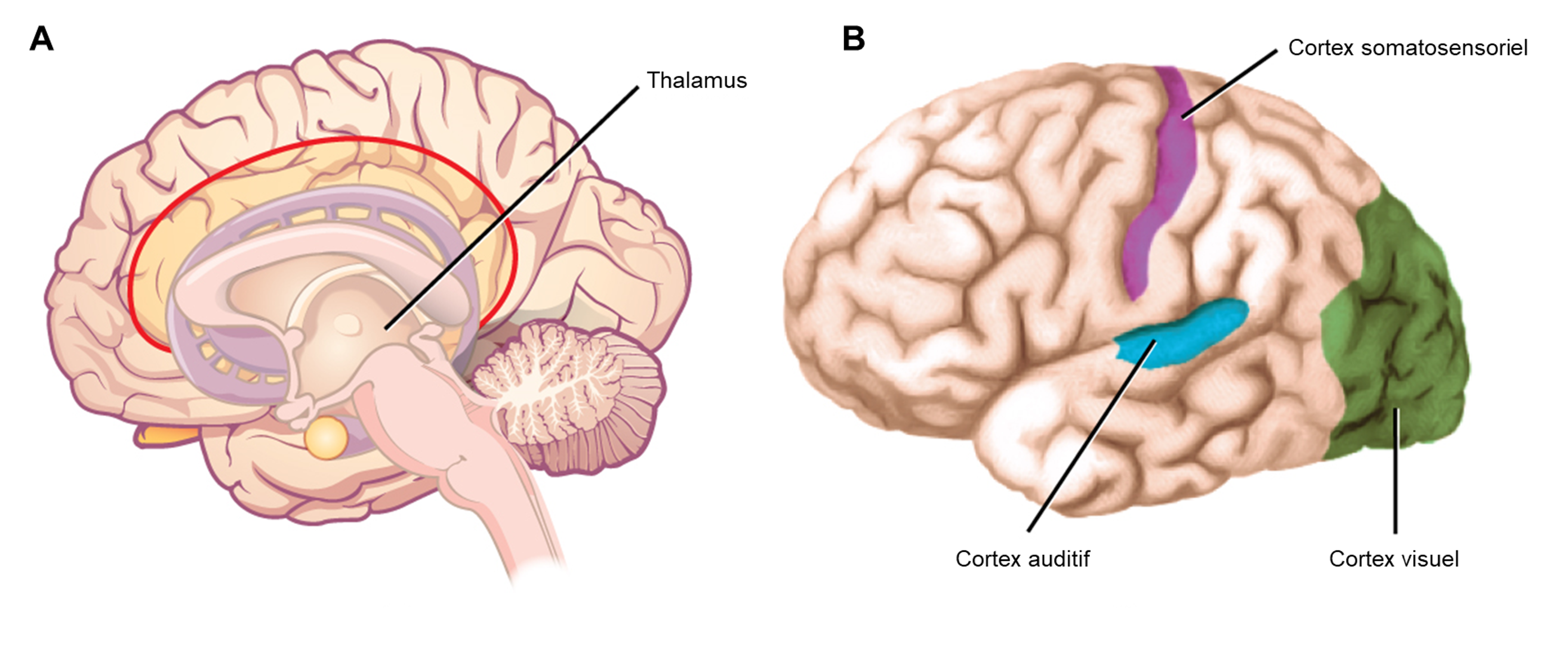
Si vous êtes intéressé par l’anatomie du système sensoriel afférent et des différentes régions du cerveau, vous pouvez les voir en détail dans ce modèle humain en trois dimensions, créé par la Society for Neuroscience américaine. Vous y trouverez des régions dont nous discuterons dans les prochains chapitres, dont les nerfs qui amènent l’information sensorielle au cerveau (nerfs crâniens optiques, olfactifs, vestibulocochléaires impliqués dans l’audition et l’équilibre) le thalamus qui reçoit les informations de différents systèmes sensoriels (vision, ouïe, toucher, proprioception, gout), le cortex visuel et auditif, le cortex somatosensoriel, ainsi que le tronc cérébral impliqué dans la respiration.
Exercices
3. Sensation et perception
Décrire la fonction sensorielle avec le terme « sensation » ou « perception » est une distinction délibérée faite par les neurophysiologistes. La sensation est l’activation des récepteurs sensoriels par un stimulus. La perception est le traitement central des stimuli sensoriels pour l’interpréter et qui implique la conscience. La perception dépend de la sensation, mais toutes les sensations ne sont pas perçues consciemment.
4. Types de cellules réceptrices : neurone afférent et cellule non neuronale
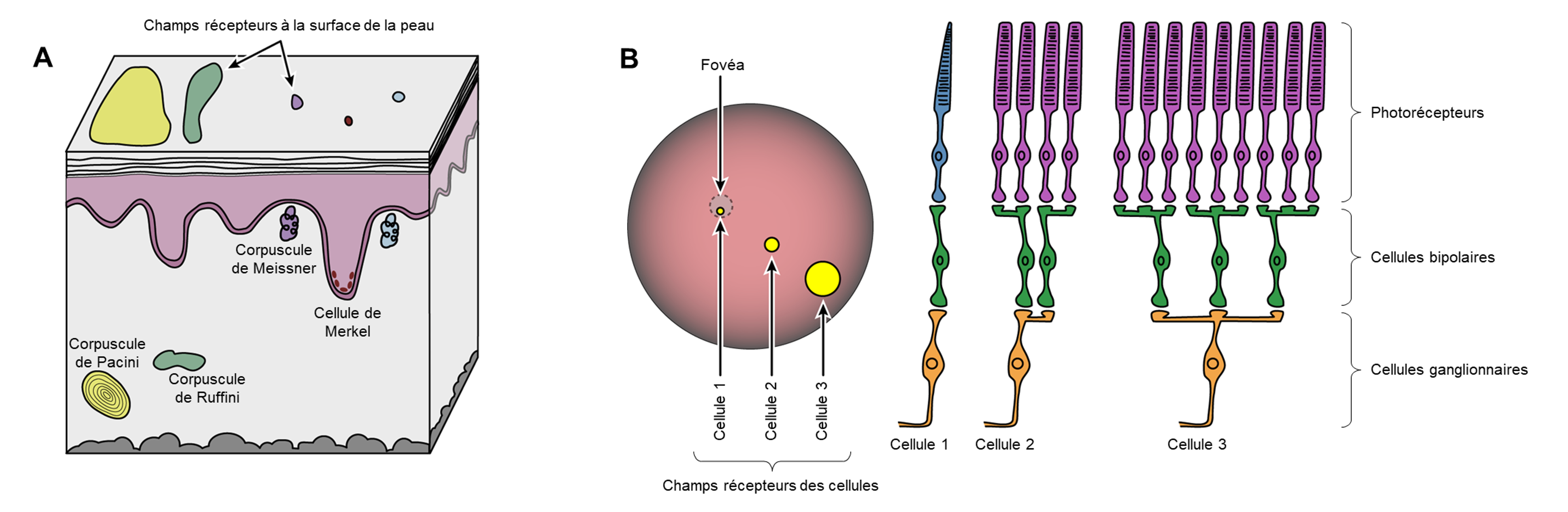
Les cellules qui interprètent les informations sur l’environnement peuvent être soit (1) un neurone afférent qui a une terminaison nerveuse (dendrites libres ou encapsulés) intégrée dans un tissu qui reçoit une sensation; (2) une cellule réceptrice spécialisée non neuronale, qui a des composants structurels distincts et qui est activée par un type spécifique de stimulus (figure 16.3). Le premier type est composé de neurones afférents (figure 16.3 A et B) qui peuvent être stimulés par plusieurs stimuli différents, montrant ainsi peu de spécificité de récepteur. Les récepteurs de la douleur et de la température dans le derme de la peau sont des exemples de neurones qui ont des terminaisons nerveuses libres. Également situés dans le derme de la peau, se trouvent des corpuscules lamellaires et tactiles, des neurones avec des terminaisons nerveuses encapsulées qui répondent à la pression et au toucher. Dans le cas du deuxième type, les cellules réceptrices spécialisées (figure 16.3C) vont détecter un stimulus spécifique et ont une grande spécificité de récepteur. Les cellules de la rétine qui répondent aux stimuli lumineux sont un exemple de cellule réceptrice spécialisée appelée un photorécepteur.
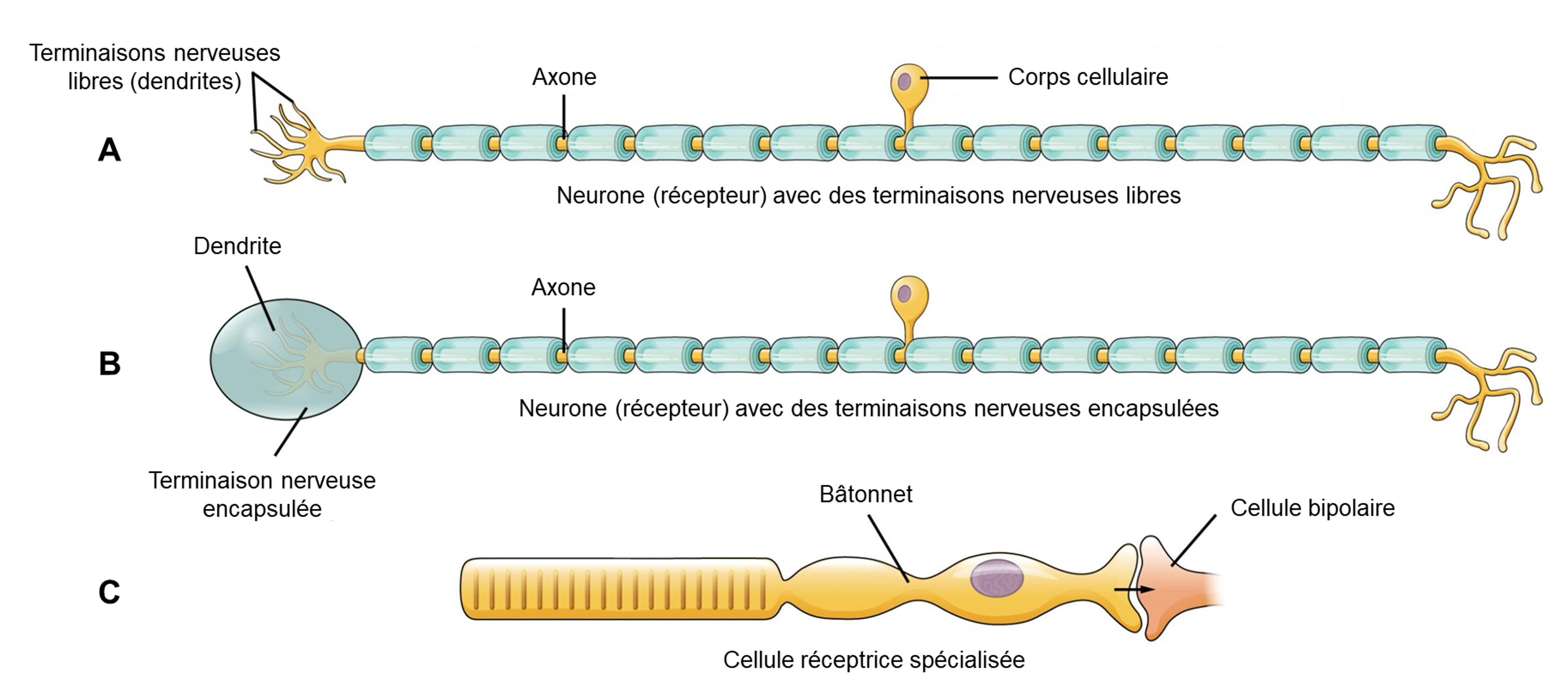
4.1 Potentiel générateur et potentiel de récepteur
Les deux grands types de systèmes cellulaires qui effectuent la transduction sensorielle ont chacune des caractéristiques particulières. Les neurones afférents produisent des potentiels gradués dans les terminaisons nerveuses libres et encapsulées qui sont appelés potentiels générateurs. Lorsqu’ils sont suffisamment forts pour atteindre le seuil, ils peuvent déclencher directement un potentiel d’action le long de l’axone du neurone sensoriel. Chez les cellules réceptrices spécialisées, les potentiels d’action déclenchés par celles-ci sont indirects et il y a deux étapes dans la transduction sensorielle. Les potentiels gradués dans les cellules réceptrices sont appelés potentiels de récepteur. Ces potentiels gradués provoquent la libération d’un neurotransmetteur dans une synapse avec un neurone sensoriel, provoquant un potentiel postsynaptique gradué. Si ce potentiel postsynaptique gradué est suffisamment fort pour atteindre le seuil, il déclenchera un potentiel d’action le long de l’axone du neurone sensoriel, qui lui transmet des informations sur le stimulus au système nerveux central. Il y a donc deux étapes dans la transmission du signal pour ce type de cellules.
4.2 Classification des récepteurs : localisation et modalité sensorielle
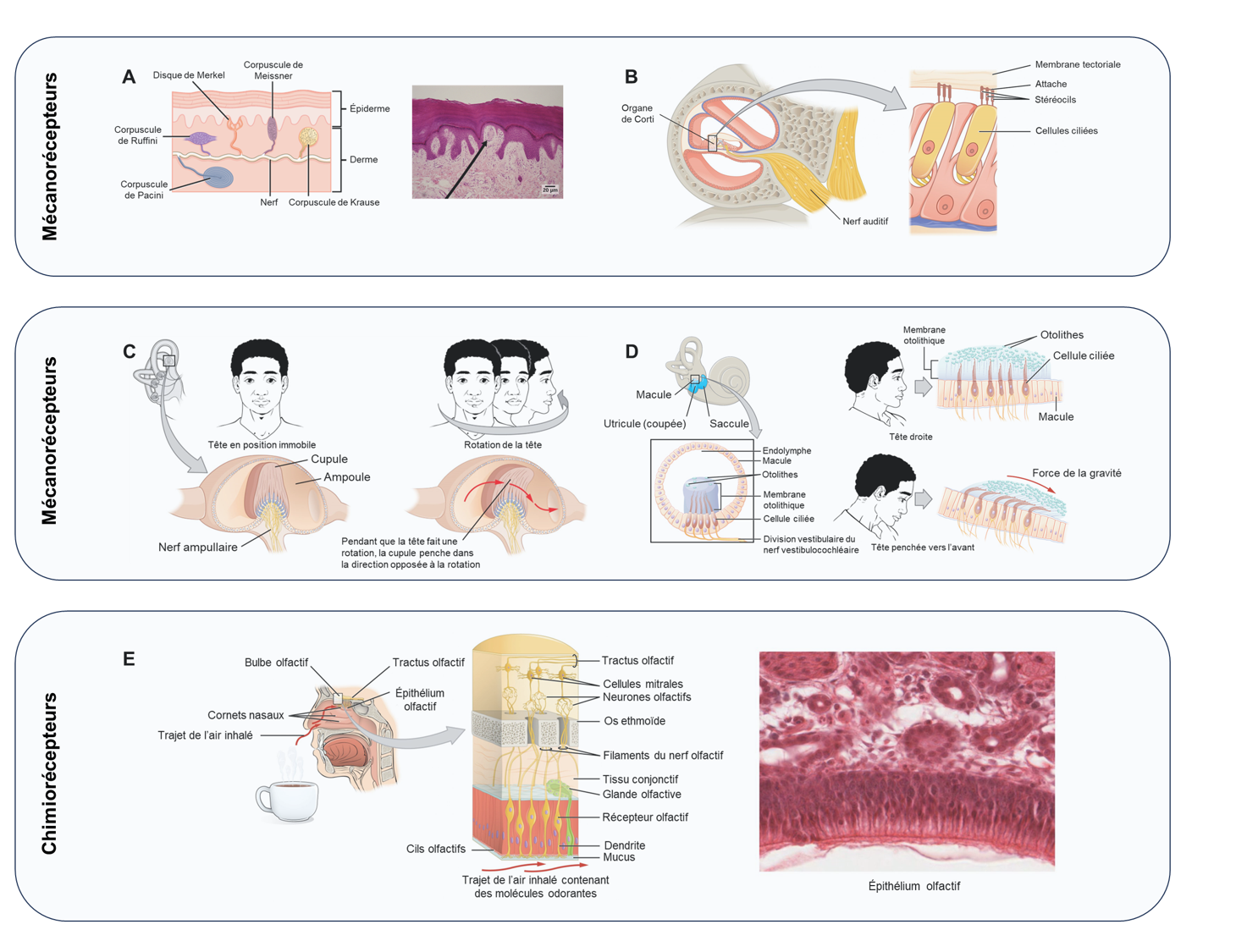
On peut classer les récepteurs en se basant sur leur localisation par rapport aux stimuli. Un extérocepteur est un récepteur situé à proximité d’un stimulus dans l’environnement externe, tel que les récepteurs somatosensoriels situés dans la peau. Un entérorécepteur est celui qui détecte les stimuli des organes et des tissus internes, tels que les récepteurs qui détectent l’augmentation de la pression artérielle dans l’aorte ou le sinus carotidien. Enfin, un propriocepteur est un récepteur situé près d’une partie mobile du corps, comme un muscle ou une capsule articulaire, qui interprète les positions des tissus lorsqu’ils bougent.
Chacun des sens est appelé une modalité sensorielle. La modalité fait référence à la manière dont l’information est encodée en une perception. Les principales modalités sensorielles peuvent être décrites sur la base de la manière dont chaque stimulus est transduit et perçu (figure 16.4). Les sens chimiques comprennent le gout et l’odorat et peuvent être détectés par des chémorécepteurs. Des chémorécepteurs permettent aussi de détecter la pression partielle d’oxygène et le pH (par la détection du CO2) ainsi que la pression osmotique dans le rein. Les osmorécepteurs répondent aux concentrations de soluté des fluides corporels. La pression, les vibrations, l’étirement musculaire et le mouvement des poils par un stimulus externe sont tous détectés par des mécanorécepteurs et sont perçus comme un toucher ou une proprioception. L’ouïe et l’équilibre sont également détectés par des mécanorécepteurs, tout comme les mouvements sont détectés par les neuromastes qui forment la ligne latérale des poissons et de certains amphibiens aquatiques, et qui sont un système de détection de la pression. Le sens qu’on appelle habituellement le toucher peut aussi comprendre une sensation qui interprète la présence de produits chimiques provenant de lésions tissulaires, ou de stimuli mécaniques intenses sous forme de douleur, détectés par un nocicepteur. Un autre stimulus physique qui a son propre type de récepteur est la température, qui est détectée par un thermorécepteur sensible aux températures supérieures (chaleur) ou inférieures (froid) à la température corporelle normale. Enfin, la catégorie des signaux électromagnétiques comprend le système visuel qui implique l’activation des photorécepteurs par la lumière du spectre du visible et des ultraviolets pour certaines espèces. On classe dans cette modalité la détection des infrarouges, des champs magnétiques et l’électrodétection que l’on retrouve chez de nombreux animaux.
Exercices
Les modalités sensorielles
5. Encodage et transmission des informations sensorielles
Quatre aspects de l’information sensorielle sont codés par les systèmes sensoriels : le type de stimulus, l’emplacement du stimulus dans le champ récepteur, l’intensité relative du stimulus et la durée. L’intégration de ces informations sensorielles commence dès que les informations sont reçues dans le système nerveux central, et le cerveau traitera ensuite les signaux entrants.
Les récepteurs sensoriels des différents sens sont très différents les uns des autres et ils sont spécialisés selon le type de stimulus qu’ils perçoivent : ils ont ce qu’on appelle une « spécificité de récepteur ». Les récepteurs transmettent des signaux nerveux par les neurones auxquels ils sont reliés, ce qui permet d’encoder le type de stimulus. Par exemple, les récepteurs tactiles ne sont pas sensibles à la lumière ou au son; ils ne sont sensibles qu’au toucher ou à la pression. Ainsi, les récepteurs tactiles transmettent des signaux sur leur propre système nerveux dédié, et l’activité électrique dans les axones des récepteurs tactiles sera donc interprétée par le cerveau comme un stimulus tactile, un toucher.
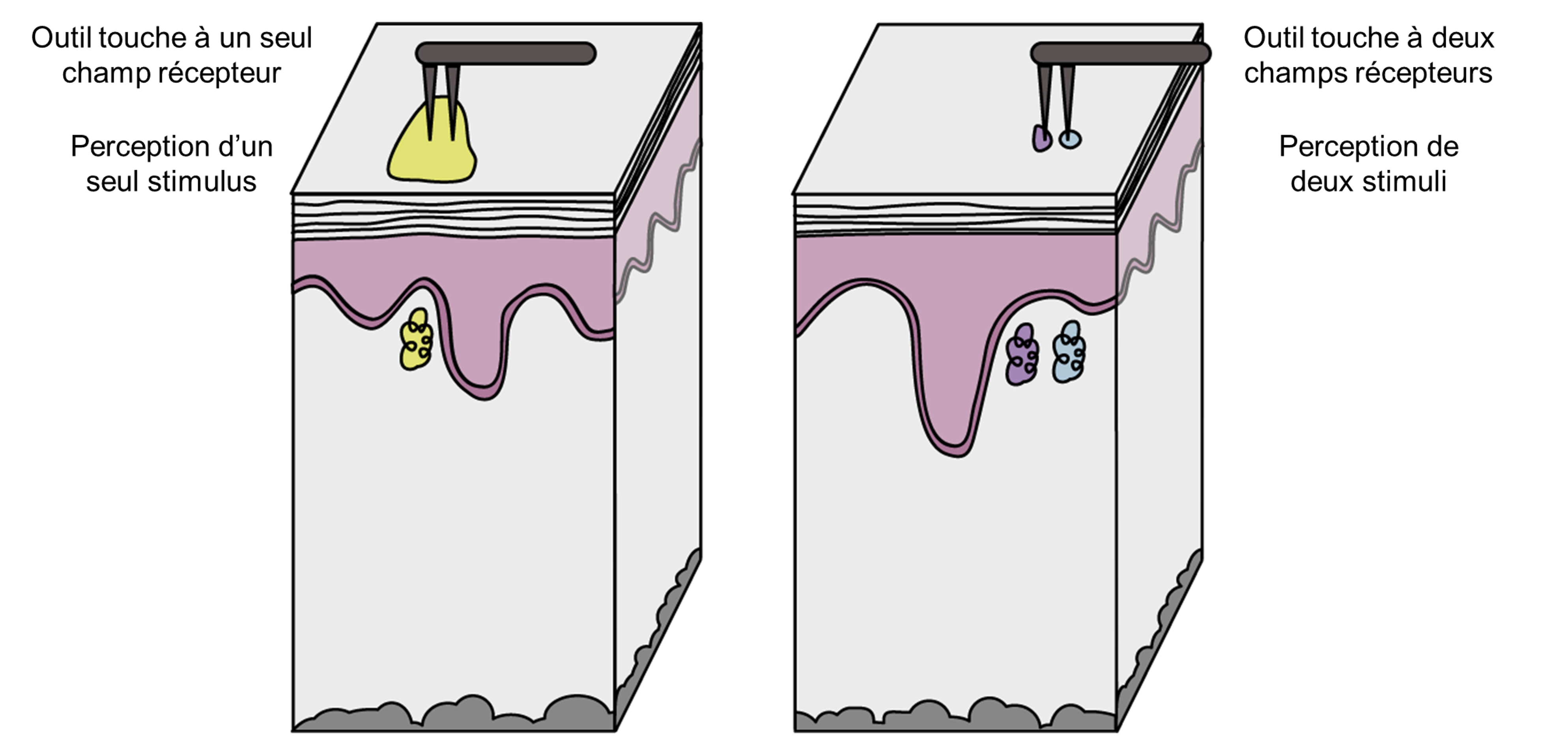
La région dans laquelle un récepteur sensoriel donné peut répondre à un stimulus est le champ récepteur. Les champs récepteurs sont plus faciles à représenter dans le système visuel et somatosensoriel. Le champ récepteur d’un neurone est la région de la rétine ou de la peau où un stimulus (lumière ou toucher) suscitera une réponse dans le neurone. Les champs récepteurs peuvent varier en taille et en forme en fonction des caractéristiques du neurone (c’est-à-dire le type, l’emplacement dans le corps, l’emplacement dans la voie). La taille des champs récepteurs est importante, car elle nous permet de localiser un stimulus sur notre corps (figure 16.5). Les champs récepteurs plus grands ne sont pas aussi précis que les champs récepteurs plus petits. Une mesure de la taille du champ récepteur d’un mécanorécepteur du toucher est la discrimination en deux points (essayez vous-même cette expérience!), qui détermine la distance minimale nécessaire entre deux stimuli pour percevoir deux points distincts sur la peau plutôt qu’un. La main a un seuil de distinction plus petit entre deux points que le dos, en raison de la taille différente des champs récepteurs.
L’intensité d’un stimulus est souvent codée dans le taux de potentiels d’action produits par le récepteur sensoriel. Ainsi, un stimulus intense produira un train plus rapide de potentiels d’action, et la réduction de l’intensité du stimulus ralentira le taux de production de potentiels d’action (figure 16.6). Une deuxième façon dont l’intensité est codée est par le nombre de récepteurs activés. Un stimulus intense pourrait initier des potentiels d’action dans un grand nombre de récepteurs adjacents, tandis qu’un stimulus moins intense pourrait stimuler moins de récepteurs. Évidemment, la durée du stimulus est encodée directement par le temps durant lequel le stimulus est perçu.
6. Intégration dans le système nerveux central et signal aux effecteurs
La section 6 est adaptée de [18].
6.1 Le thalamus et le cortex cérébral
Dans le système nerveux somatique des humains, le thalamus est un relai important de communication. L’apport sensoriel au thalamus provient de la plupart des sens spécifiques et des voies somatosensorielles. Chaque système sensoriel est relayé par un noyau particulier dans le thalamus. Le thalamus est un point de transfert nécessaire pour la plupart des voies sensorielles qui atteignent le cortex cérébral, où commence la perception sensorielle consciente. La seule exception à cette règle est le système olfactif. Les axones du tractus olfactif du bulbe olfactif se projettent directement vers le cortex cérébral.
6.2 Topographie sensorielle
Un aspect fascinant de l’organisation du système nerveux sensoriel est que de nombreux axones sensoriels sont positionnés de la même manière que leurs cellules réceptrices correspondantes dans le corps. Cela permet au cerveau d’identifier la position d’un stimulus sur la base de quelles cellules réceptrices envoient des informations. Le cortex cérébral entretient également cette topographie sensorielle dans les zones particulières du cortex qui correspondent à la position des cellules réceptrices. La représentation spatiale des cellules réceptrices est donc maintenue dans le cerveau et les neurones qui sont un à côté de l’autre dans le cortex reçoivent des informations des récepteurs sensoriels qui sont un à côté de l’autre, par exemple sur la peau. Le cortex somatosensoriel fournit en effet un exemple dans lequel, essentiellement, les emplacements des récepteurs somatosensoriels dans le corps sont cartographiés sur le cortex somatosensoriel.
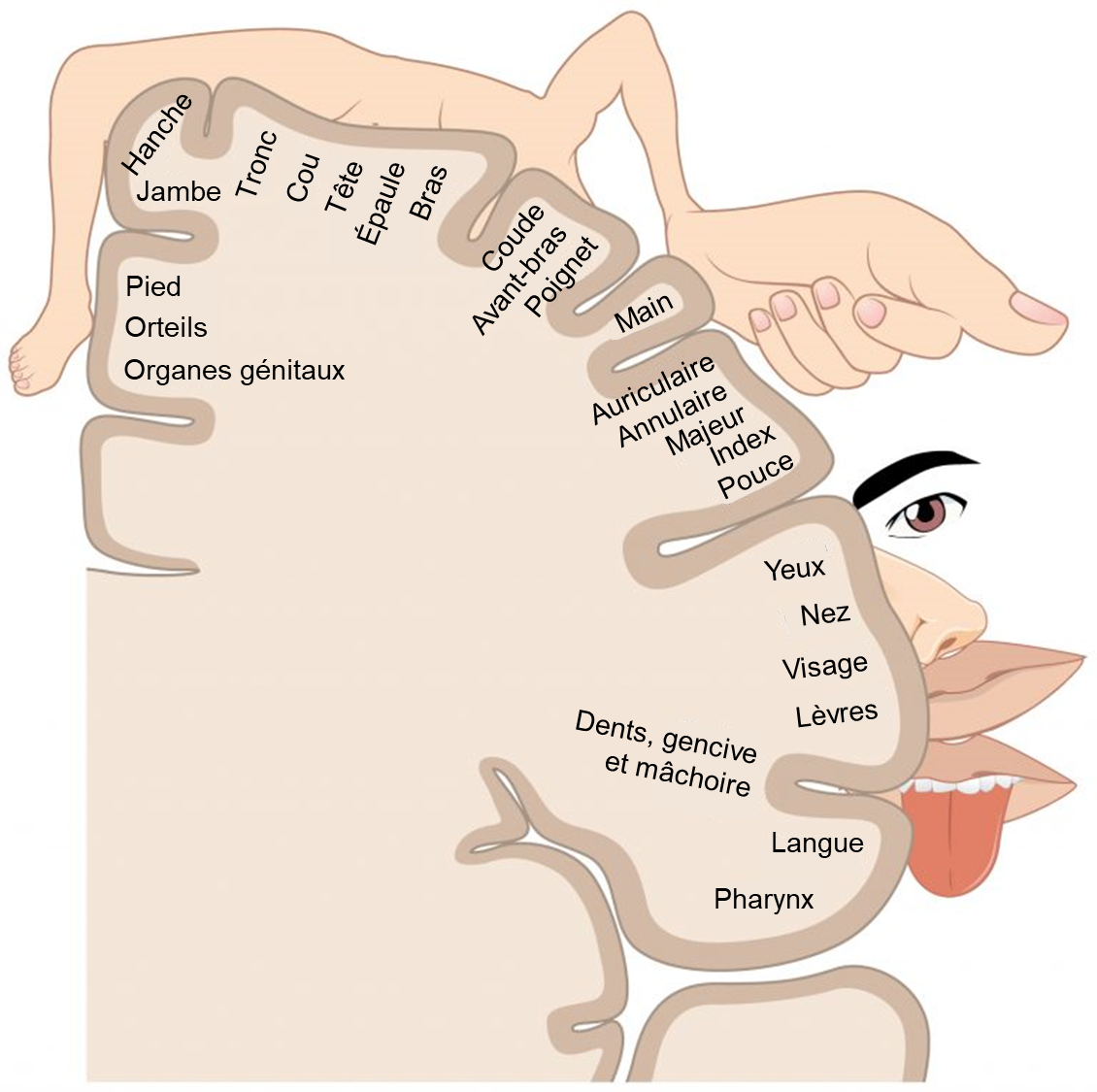
Cette cartographie est souvent représentée à l’aide d’un « homoncule » sensoriel. Le terme homonculus vient du mot latin signifiant « petit homme » et fait référence à une carte du corps humain posée sur une carte du cortex cérébral (figure 16.7). Dans le cortex somatosensoriel, les organes génitaux externes, les pieds et le bas des jambes sont représentés ensemble. La carte corporelle se poursuit à travers les cuisses, les hanches, le tronc, les épaules, les bras et les mains. La tête et le visage sont juste latéraux aux doigts dans le cortex. Cette représentation dans le cortex s’appelle une carte corticale. Ce type de maintien de l’information spatiale des récepteurs jusqu’au cortex se retrouve aussi dans le système visuel. Il est intéressant de noter que l’idée de carte corticale a été développée par le Dr. Wilder Penfield, un québécois d’adoption qui travaillait à l’université McGill.
6.3 La carte corticale est proportionnelle à la quantité d’information du système sensoriel et est plastique
En plus de l’existence de cette carte corticale, une caractéristique fascinante est que cette correspondance ne donne pas lieu à une version parfaitement miniature du corps, mais exagère plutôt les zones les plus sensibles du corps, comme les doigts et le bas du visage chez l’humain. Ces zones sont représentées par plus de neurones et donc l’information est plus précise. Il a aussi été proposé que dans certains cas, le même nombre de neurones résulte en une plus grande représentation dans le cortex si cette information est importante pour la performance de l’animal. Les zones moins sensibles du corps, telles que les épaules et le dos chez l’humain, sont cartographiées sur des zones plus petites du cortex.
Il est important de noter que cette carte corticale est plastique à l’intérieur de la vie d’un organisme et la taille de différentes régions peut changer avec l’expérience ou des changements au système sensoriel (perte d’un membre, d’un sens spécifique, apprentissage moteur). Des neurobiologistes ont par exemple placé des rats en cage standard et ont cartographié la région du cortex qui recevait les informations sensorielles de vibrisses individuelles de leurs moustaches. Ils les ont transférés dans un milieu imitant leur habitat naturel qui incluait des conspécifiques et la possibilité de creuser des tunnels et ont observé des changements dans la carte corticale après 4 semaines. Après avoir été utilisée dans un environnement procurant plus d’information sensorielle, chaque vibrisse de la moustache avait un champ récepteur plus petit et les champs récepteurs des 3 vibrisses étudiés n’étaient plus superposés, indiquant que le cerveau interprétait maintenant les signaux de ces vibrisses de façon individuelle et plus précise. Ceci démontre une plasticité importante même chez les mammifères adultes qui permet un raffinement de la carte corticale selon l’expérience [20]. La carte corticale répond donc aux informations sensorielles reçues.
6.4 La carte corticale varie entre les espèces
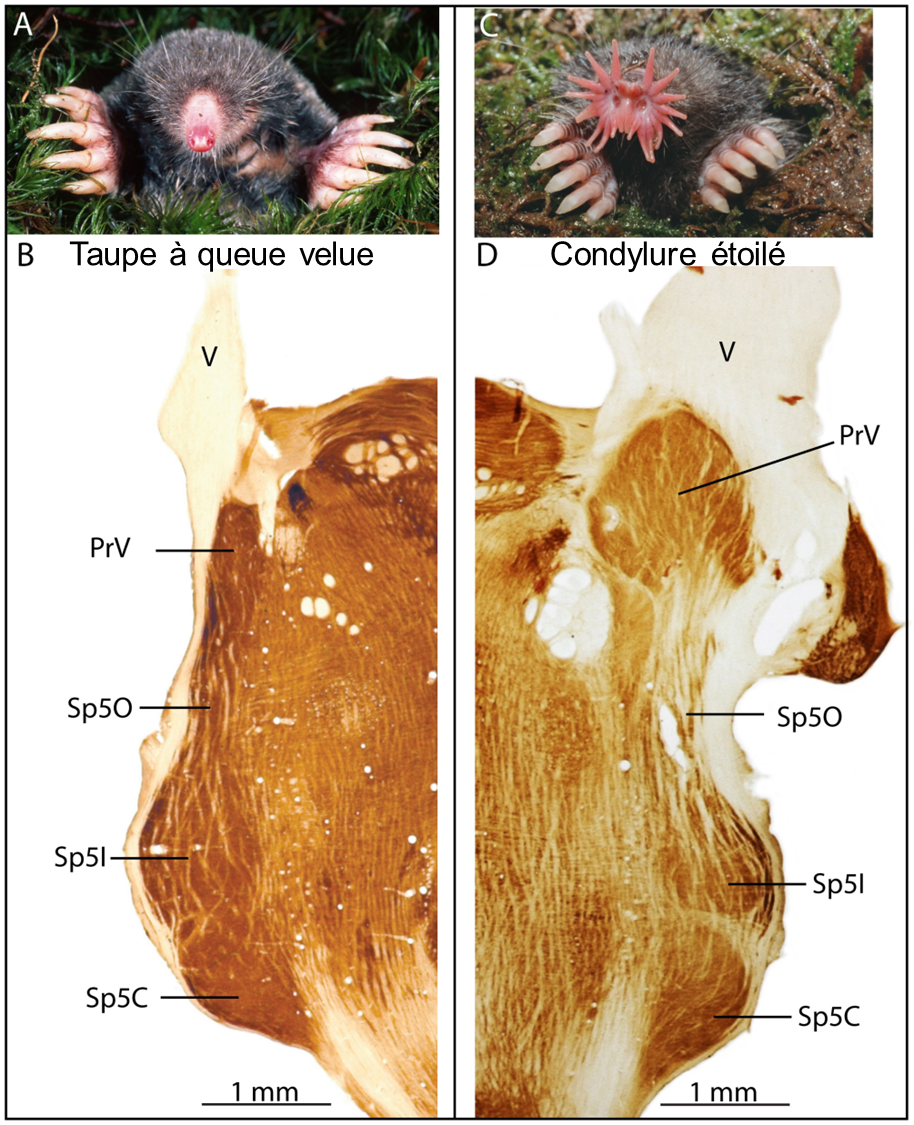
On peut cartographier la représentation du système sensoriel dans le cortex de différentes espèces et obtenir des cartes corticales différentes, si les régions du corps qui ont plus de récepteurs sensoriels ne sont pas les mêmes entre ces espèces. Ceci représente l’évolution des systèmes sensoriels de ces différentes espèces. Par exemple, des neurobiologistes ont comparé deux espèces de taupes qui sont similaires excepté pour une structure sensorielle. La taupe à queue velue a un nez typique d’une taupe alors que le condylure étoilé a une structure mécanosensorielle en forme d’étoile au bout du museau (figure 16.8 A et C). Les neurobiologistes se sont intéressés au noyau sensoriel trijumeau principal (PrV sur la figure 16.8 B et D) dans le tronc cérébral des condylures étoilés. Le PrV reçoit les informations du nerf trijumeau qui apporte des signaux du toucher et se projette via le thalamus vers le cortex somatosensoriel. Leurs résultats révèlent que le PrV est massivement étendu par rapport au PrV chez les rongeurs ou les taupes sans nez étoilé (figure 16.8). La carte corticale a évolué entre les deux espèces.
Cette vidéo présente la topographie sensorielle.
Aubin-Horth, N. (2024). Écophysiologie évolutive: la topographie sensorielle. [Vidéo]. Youtube. CC-BY. https://youtu.be/Gw22N88jMFY?si=raAk67seQS8htQ1K
Vous trouverez la transcription de l’audio de cette capsule en cliquant sur ce lien: Transcription en format word.
7. Réponses motrices
La section 7 est adaptée de [18]
La caractéristique déterminante du système nerveux somatique est qu’il contrôle les muscles squelettiques. Les sens somatiques informent le système nerveux sur l’environnement extérieur et la réponse à ces informations se fait par le biais de mouvements musculaires volontaires. Le terme « volontaire » suggère qu’il existe une décision consciente d’effectuer un mouvement. Cependant, certains aspects du système somatique sollicitent les muscles volontaires sans contrôle conscient. Un exemple est la capacité de notre respiration à passer à un contrôle inconscient pendant que nous sommes concentrés sur une autre tâche. Cependant, les muscles responsables du processus de base de la respiration sont également utilisés pour la parole, qui est entièrement volontaire.
Les fonctions motrices sont largement contrôlées par le lobe frontal. Les régions les plus antérieures du lobe frontal — les zones préfrontales — sont importantes pour les fonctions exécutives, qui sont les fonctions cognitives qui conduisent à des comportements orientés vers un objectif. Ces processus cognitifs supérieurs incluent la mémoire de travail, appelée « bloc-notes mental », qui peut aider à organiser et à représenter des informations qui ne se trouvent pas dans l’environnement immédiat. Le lobe préfrontal est responsable d’aspects de l’attention, tels que l’inhibition des pensées et des actions distrayantes afin qu’une personne puisse se concentrer sur un objectif et orienter son comportement vers l’atteinte de cet objectif.
Les régions restantes du lobe frontal sont les régions du cortex qui produisent le mouvement. Les zones préfrontales se projettent dans le cortex moteur secondaire, qui comprend le cortex prémoteur et l’aire motrice supplémentaire, deux régions importantes qui aident à la planification et à la coordination des mouvements. Le cortex prémoteur aide à contrôler les mouvements des muscles centraux pour maintenir la posture pendant le mouvement, tandis que l’aire motrice supplémentaire serait responsable de la planification et de la coordination des mouvements, ainsi que des mouvements séquentiels basés sur une expérience antérieure (c’est-à-dire des mouvements appris). Les neurones de ces zones sont les plus actifs avant l’initiation du mouvement. Finalement, le cortex moteur primaire reçoit des informations de plusieurs zones qui aident à planifier le mouvement. La sortie motrice du cortex moteur primaire descend dans le tronc cérébral et dans la moelle épinière via les neurones moteurs pour stimuler la contraction des muscles squelettiques.
Cas appliqué
Texte adapté de [22].
Les cerveaux sont composés de plusieurs régions dont la taille varie considérablement selon l’espèce de vertébré et qui sont associées à des fonctions et à des comportements particuliers. La taille des régions cérébrales évolue généralement de manière allométrique avec la taille du cerveau. Habituellement, plus un cerveau augmente en taille, plus chaque région augmente en taille.
Cependant, des changements en mosaïque dans la taille des régions cérébrales, indépendants de la taille du cerveau, ont été observés dans plusieurs lignées et pourraient être liés à l’évolution de nouveaux comportements. Ces changements reflèteraient les pressions de sélection sur les traits associés à ces régions. Les biologistes supposent que l’évolution cérébrale en mosaïque des principales régions du cerveau se produit parallèlement à des innovations comportementales qui ouvrent de nouvelles niches, car les changements en mosaïque sont plus susceptibles de contribuer à des différences majeures dans la fonction cérébrale.
Les poissons faiblement électriques sont excellents pour tester si l’évolution du cerveau en mosaïque se produit avec une nouveauté comportementale : ces poissons ont développé de nouveaux systèmes électrosensoriels actifs sur le plan comportemental avec plusieurs innovations neuronales, ce qui a probablement abouti à une forte sélection pour les capacités de traitement électrosensoriel. De plus, plusieurs lignées de poissons ont développé indépendamment des systèmes électrosensoriels similaires, un cas d’évolution convergente.
Des études ont montré que les poissons africains faiblement électriques de la famille des Mormyridés développent des cerveaux extrêmement gros par rapport à leur taille corporelle et montrent une augmentation en mosaïque de la taille du cervelet et du tronc cérébral par rapport à d’autres espèces proches évolutivement mais qui ne sont pas électriques. Ces augmentations de la taille de certaines régions spécifiques se sont produites parallèlement à l’évolution d’un système électrosensoriel actif. Ils ont la capacité d’électrogenèse, soit la production d’électricité, ainsi que la capacité d’électroréception. Cependant, comme il ne s’agit que d’une seule lignée, il est impossible de déterminer si ces changements de mosaïque sont associés à l’évolution de ce système électrosensoriel ou à d’autres phénotypes qui divergent entre les Mormyridés et leurs espèces proches. Les biologistes évolutifs ont résolu ce problème en étudiant les poissons faiblement électriques de l’ordre des Gymnotiformes et les poissons-chats faiblement électriques d’Amérique du Sud (de l’ordre des siluriformes), qui ont évolué des systèmes électrosensoriels de façon indépendante des mormyroïdes (figure 16.9).
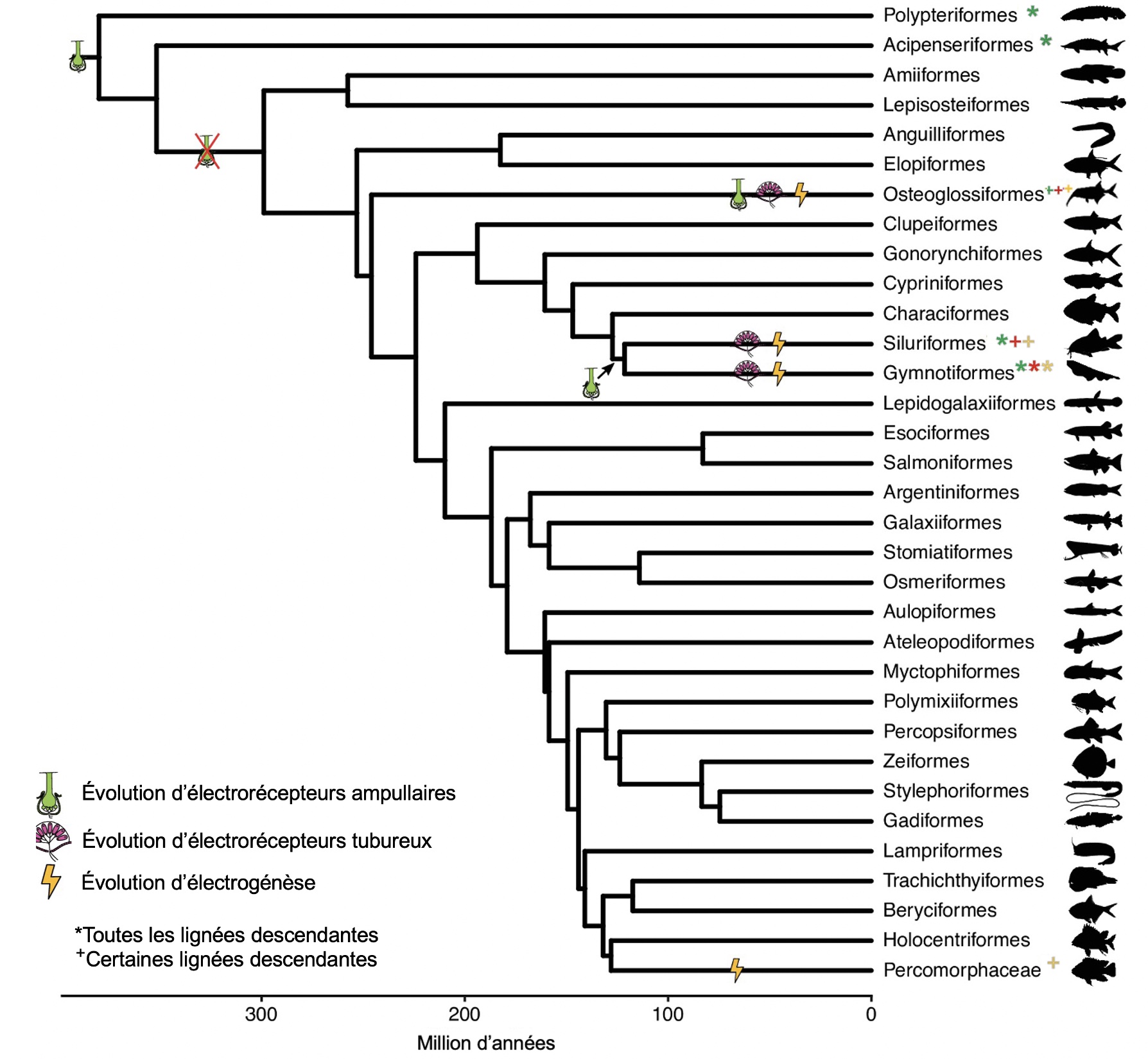
Si l’augmentation de la taille de certaines régions du cerveau des mormyridés est liée à l’évolution d’un système électrosensoriel, nous devrions observer des changements en mosaïque similaires chez les gymnotiformes et les siluriformes. Les biologistes ont quantifié la taille des régions cérébrales pour plusieurs espèces électrogènes, électroréceptives et non électrosensibles. Ils ont utilisé la microtomographie, une technique qui utilise des rayons X pour créer un modèle en trois dimensions d’un objet. Ils ont constaté une augmentation de la taille du cervelet dans les trois lignées produisant de l’électricité par rapport aux lignées non électriques, ainsi qu’une augmentation de la mosaïque dans le tronc cérébral associée à l’évolution de l’électrogenèse et du type d’électrorécepteur (figure 16.10). Le traitement de l’information électrosensorielle qui arrive des récepteurs a lieu dans le tronc cérébral. Le tronc cérébral est également impliqué dans la génération d’électricité, ce qui suggère que le traitement électrosensoriel et le contrôle électromoteur sont associés aux changements évolutifs dans la taille relative de ces régions. Les neurobiologistes ont comme prémisse que cette augmentation en taille de la région est associée à une augmentation du nombre de neurones, ce qui augmenterait la sensibilité du système sensoriel et la capacité de traiter de l’information. Cependant, ils n’excluent pas que la taille des neurones puisse aussi avoir évolué.
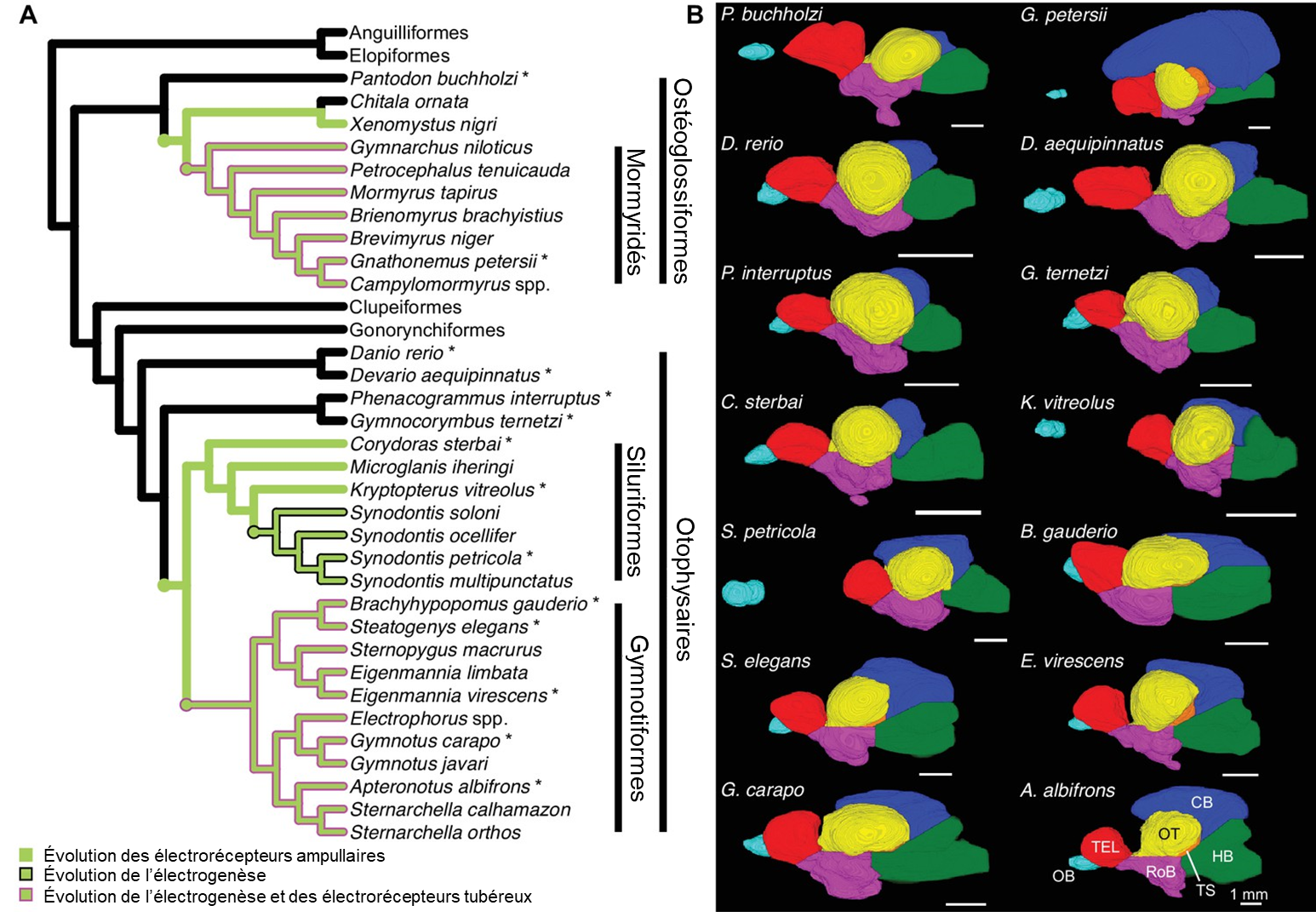
Ces résultats montrent que l’évolution de nouveaux systèmes électrosensoriels est associée de manière répétée et indépendante à des changements dans la taille des principales régions cérébrales individuelles qui sont impliquées dans ce système sensoriel et moteur, indépendamment de la taille du cerveau, ce qui suggère que la sélection peut avoir un impact sur la composition structurelle du cerveau afin de favoriser des régions spécifiques impliquées dans de nouveaux comportements.
Résumé
Une caractéristique commune des différentes composantes du système sensoriel est leur fonction de transduction sensorielle, qui inclut les étapes de réception, transduction et perception. Il y a deux types de cellules réceptrices, soit les neurones afférents et les cellules réceptrices spécialisées non neuronales. On peut classer les récepteurs sensoriels selon leur localisation et selon leur modalité sensorielle. Les caractéristiques d’un stimulus (le type de stimulus, l’emplacement du stimulus dans le champ récepteur, l’intensité relative du stimulus et sa durée) sont encodées par le système sensoriel et le système nerveux central. L’information sensorielle est intégrée par le système nerveux central en passant la plupart du temps par le thalamus pour être ensuite dirigée vers le cortex cérébral. La représentation spatiale des cellules réceptrices est maintenue dans le cerveau, formant une carte corticale. Celle-ci est proportionnelle à la quantité d’information du système sensoriel, est plastique et peut varier entre les espèces. Les informations sensorielles une fois intégrées peuvent mener à une réponse motrice, contrôlée par le lobe frontal.
Activité de révision
Exercices
Révision sur le système sensoriel
Bibliographie
- Biga, L.M., S. Bronson, S. Dawson, A. Harwell, R. Hopkins, J. Kaufmann, M. LeMaster, P. Matern, K. Morrison-Graham, K. Oja, D. Quick, J. Runyeon, O. OERU, and OpenStax (2019). Sensory Receptors, in Anatomy & Physiology [CC-BY-SA]. Oregon State University. https://open.oregonstate.education/aandp/chapter/13-1-sensory-receptors/.
- Molnar, C. and J. Gair (2015). Sensory Processes, in Concepts of Biology – 1st Canadian Edition [CC-BY]. BC Campus. https://opentextbc.ca/biology/chapter/17-1-sensory-processes/.
- Henley, C. (2021). Touch: The Skin, in Foundations of Neuroscience [CC-BY-NC-SA]. Libraries Michigan State University. https://openbooks.lib.msu.edu/neuroscience/chapter/touch-the-skin/.
- Molnar, C. and J. Gair (2015). Mechanosensitive ion channels are gated ion channels that respond to mechanical deformation of the plasma membrane. [CC-BY]. https://opentextbc.ca/biology/chapter/17-1-sensory-processes/.
- Henley, C. (2021). Receptive Field Activation [CC-BY-NC-SA]. https://openbooks.lib.msu.edu/neuroscience/chapter/touch-the-skin/.
- Molnar, C. and J. Gair (2015). In humans, with the exception of olfaction, all sensory signals are routed from the thalamus to final processing regions in the cortex of the brain. [CC-BY]. https://opentextbc.ca/biology/chapter/17-1-sensory-processes/.
- Shigeru23 (2011). Structure of sensory system 4 models with English tags [CC-BY-SA]. https://commons.wikimedia.org/wiki/File:Structure_of_sensory_system_(4_models)_E.PNG.
- Betts, J.G., P. Desaix, E. Johnson, J.E. Johnson, O. Korol, D. Kruse, B. Poe, J.A. Wise, M. Womble, and K.A. Young (2022). Receptor Classification by Cell Type. OpenStax. [CC-BY]. https://openstax.org/books/anatomy-and-physiology-2e/pages/14-1-sensory-perception.
- Molnar, C. and J. Gair (2015). Four of the primary mechanoreceptors in human skin. [CC-BY]. https://opentextbc.ca/biology/chapter/17-2-somatosensation/.
- Molnar, C. and J. Gair (2015). Meissner corpuscles in the fingertips. [CC-BY]. https://opentextbc.ca/biology/chapter/17-2-somatosensation/.
- Betts, J.G., P. Desaix, E. Johnson, J.E. Johnson, O. Korol, D. Kruse, B. Poe, J.A. Wise, M. Womble, and K.A. Young (2022). The Hair Cell. OpenStax. [CC-BY]. https://openstax.org/books/anatomy-and-physiology-2e/pages/14-1-sensory-perception.
- Betts, J.G., P. Desaix, E. Johnson, J.E. Johnson, O. Korol, D. Kruse, B. Poe, J.A. Wise, M. Womble, and K.A. Young (2022). Rotational Coding by Semicircular Canals. OpenStax. [CC-BY]. https://openstax.org/books/anatomy-and-physiology-2e/pages/14-1-sensory-perception.
- Betts, J.G., P. Desaix, E. Johnson, J.E. Johnson, O. Korol, D. Kruse, B. Poe, J.A. Wise, M. Womble, and K.A. Young (2022). Linear Acceleration Coding by Maculae. OpenStax. [CC-BY]. https://openstax.org/books/anatomy-and-physiology-2e/pages/14-1-sensory-perception.
- Betts, J.G., P. Desaix, E. Johnson, J.E. Johnson, O. Korol, D. Kruse, B. Poe, J.A. Wise, M. Womble, and K.A. Young (2022). The Olfactory System. OpenStax. [CC-BY]. https://openstax.org/books/anatomy-and-physiology-2e/pages/14-1-sensory-perception.
- Henley, C. (2021). Two-Point Discrimination [CC-BY-NC-SA]. https://openbooks.lib.msu.edu/neuroscience/chapter/touch-the-skin/.
- Henley, C. (2021). Mechanoreceptor Receptive Fields [CC-BY-NC-SA]. https://openbooks.lib.msu.edu/neuroscience/chapter/touch-the-skin/.
- Henley, C. (2021). Retinal Receptive Field [CC-BY-NC-SA]. https://openbooks.lib.msu.edu/neuroscience/chapter/vision-the-retina/.
- Biga, L.M., S. Bronson, S. Dawson, A. Harwell, R. Hopkins, J. Kaufmann, M. LeMaster, P. Matern, K. Morrison-Graham, K. Oja, D. Quick, J. Runyeon, O. OERU, and OpenStax (2019). Sensory and Motor Pathways, in Anatomy & Physiology [CC-BY-SA]. Oregon State University. https://open.oregonstate.education/aandp/chapter/14-5-sensory-and-motor-pathways/.
- Betts, J.G., P. Desaix, E. Johnson, J.E. Johnson, O. Korol, D. Kruse, B. Poe, J.A. Wise, M. Womble, and K.A. Young (2022). The Sensory Homunculus. OpenStax [CC-BY]. https://openstax.org/books/anatomy-and-physiology-2e/pages/14-2-central-processing.
- Polley, D.B., E. Kvašňák, and R.D. Frostig (2004). Naturalistic experience transforms sensory maps in the adult cortex of caged animals Nature. 429(6987): p. 67-71. https://doi.org/10.1038/nature02469.
- Catania, K.C., D.B. Leitch, and D. Gauthier (2011). A Star in the Brainstem Reveals the First Step of Cortical Magnification [CC-BY]. PLOS ONE. 6(7): p. e22406. https://doi.org/10.1371/journal.pone.0022406.
- Schumacher, E.L. and B.A. Carlson (2022). Convergent mosaic brain evolution is associated with the evolution of novel electrosensory systems in teleost fishes [CC-BY]. eLife. 11. https://doi.org/10.7554/eLife.74159.

_E.PNG){kind=link}